

Induced expression of expanded CGG RNA causes mitochondrial dysfunction in vivo
- PMID: 25486200
- PMCID: PMC4614669
- DOI: 10.4161/15384101.2014.943112
Induced expression of expanded CGG RNA causes mitochondrial dysfunction in vivo
Abstract
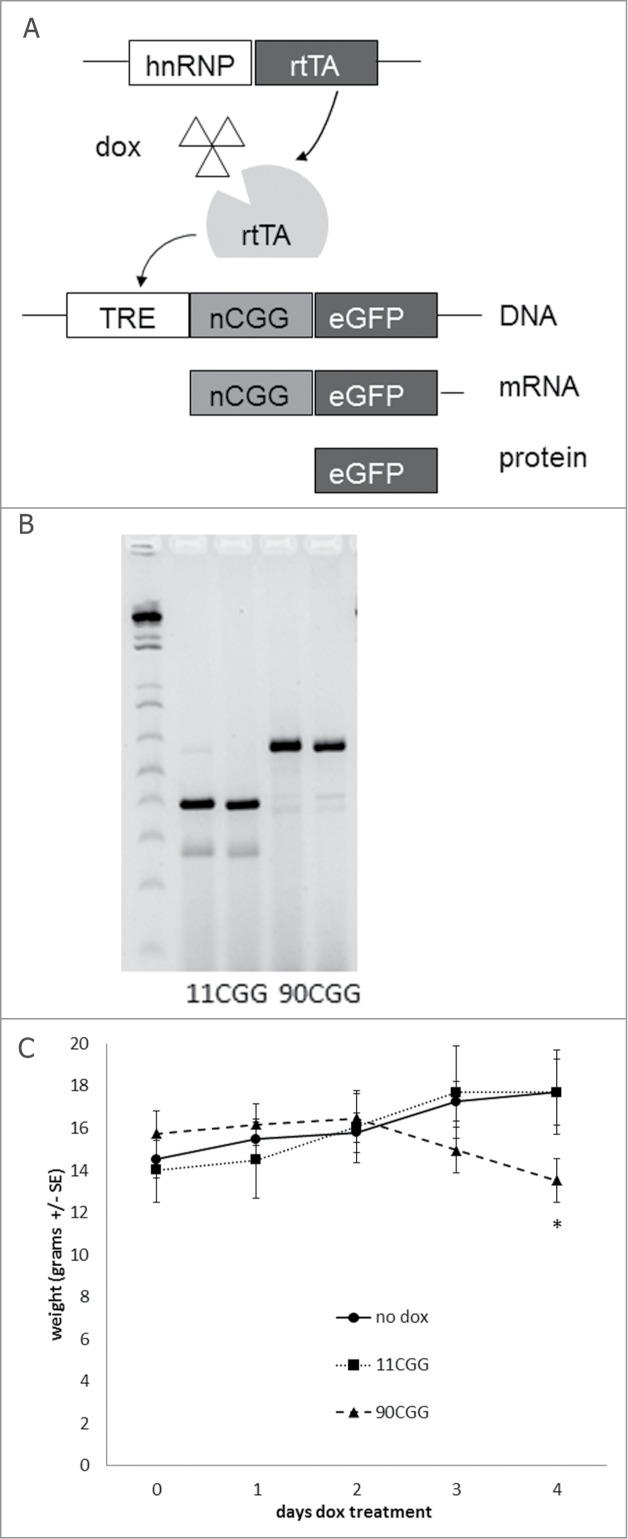
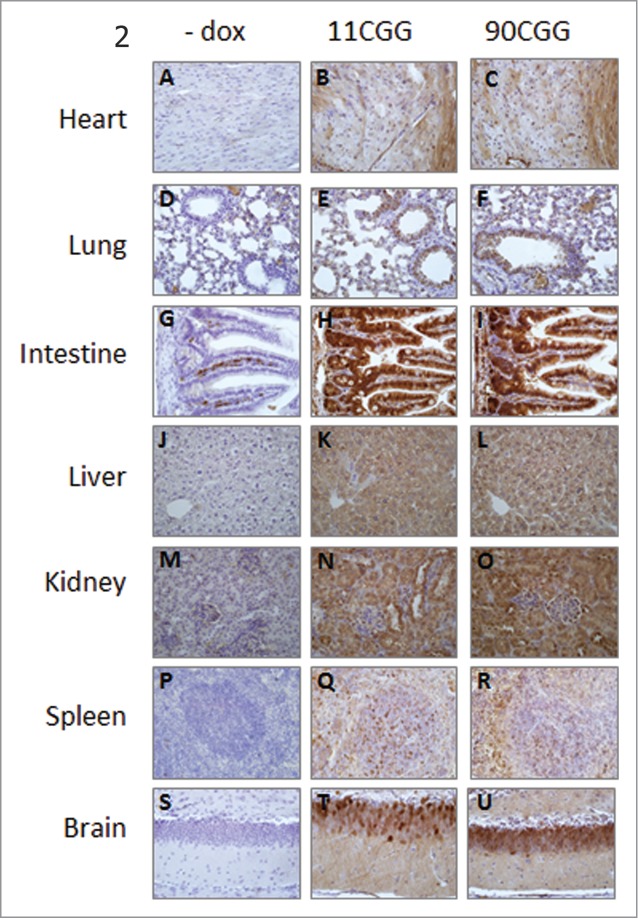
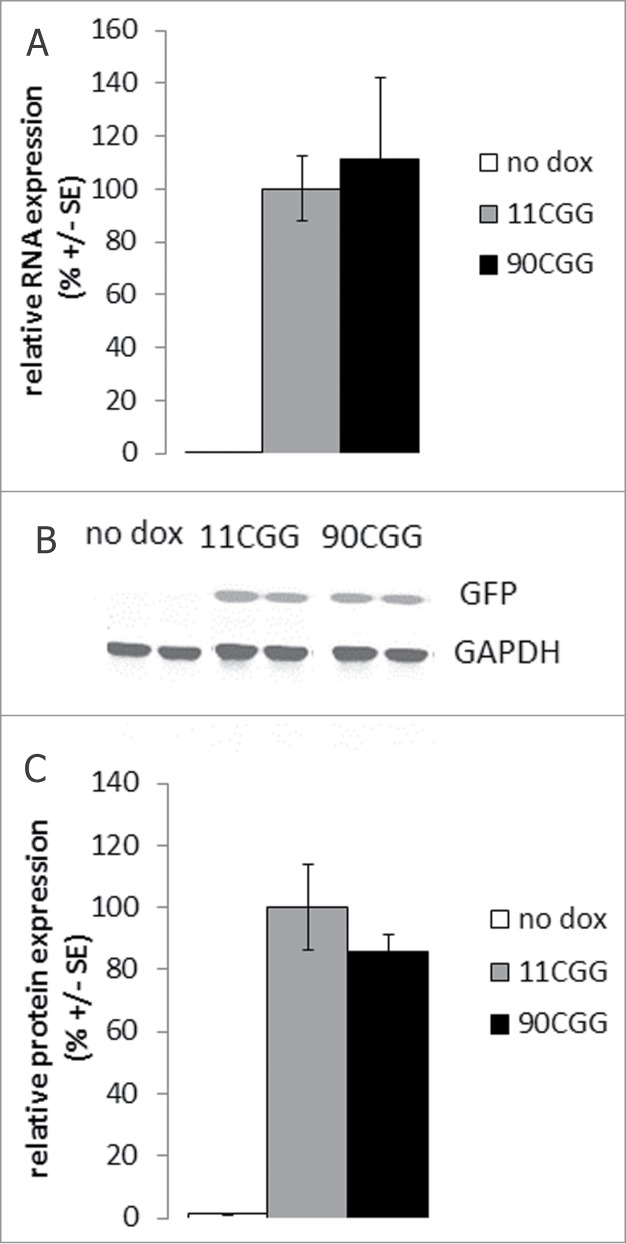
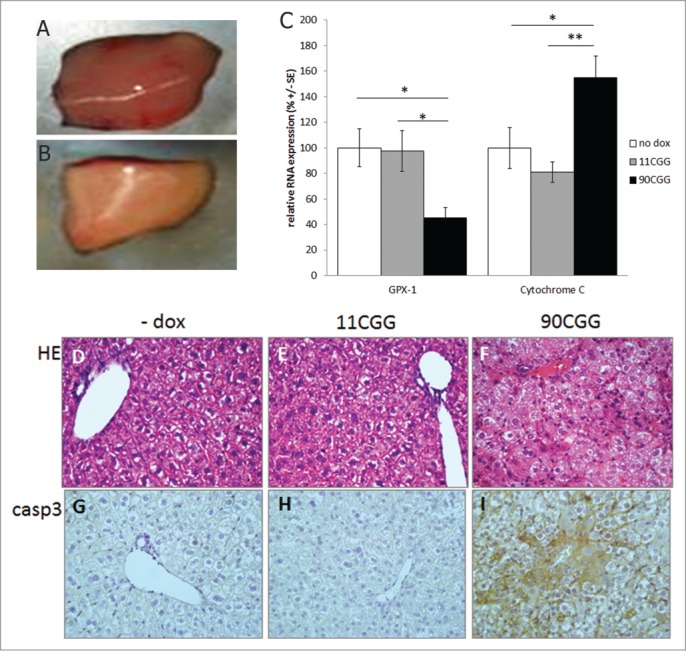
Fragile X-associated tremor/ataxia syndrome (FXTAS) is a late-onset neurodegenerative disorder affecting carriers of premutation forms of the FMR1 gene, resulting in a progressive development of tremor, ataxia and neuropsychological problems. The disease is caused by an expanded CGG repeat in the FMR1 gene, leading to an RNA gain-of-function toxicity mechanism. In order to study the pathogenesis of FXTAS, new inducible transgenic mouse models have been developed that expresses either 11CGGs or 90CGGs at the RNA level under control of a Tet-On promoter. When bred to an hnRNP-rtTA driver line, doxycycline (dox) induced expression of the transgene could be found in almost all tissues. Dox exposure resulted in loss of weight and death within 5 d for the 90CGG RNA expressing mice. Immunohistochemical examination of tissues of these mice revealed steatosis and apoptosis in the liver. Decreased expression of GPX1 and increased expression of cytochrome C is found. These effects were not seen in mice expressing a normal sized 11CGG repeat. In conclusion, we were able to show in vivo that expression of an expanded CGG-repeat rather than overexpression of a normal CGG-repeat causes pathology. In addition, we have shown that expanded CGG RNA expression can cause mitochondrial dysfunction by regulating expression levels of several markers. Although FTXAS patients do not display liver abnormalities, our findings contribute to understanding of the molecular mechanisms underlying toxicity of CGG repeat RNA expression in an animal model. In addition, the dox inducible mouse lines offer new opportunities to study therapeutic interventions for FXTAS.
Keywords: CGG repeat; FXTAS; FXTAS, Fragile X-associated tremor/ataxia syndrome; RNA gain-of-function; TRE, Tet Responsive Element; Tet-On; apoptosis; caspase 3; cytochrome C; dox, doxycycline; eGFP, enhanced green fluorescent protein; gpx, gluthation peroxidase; gpx-1; inducible mouse model; mitochondria; rtTA, reverse tetracycline transactivator.
Figures
Comment in
-
FXTAS: size does matter!Cell Cycle. 2014;13(21):3319. doi: 10.4161/15384101.2014.972920. Cell Cycle. 2014. PMID: 25485575 Free PMC article. No abstract available.
References
-
- Willemsen R, Levenga J, Oostra B. CGG repeat in the FMR1 gene: size matters. ClinGenet 2011; 80:214–25; PMID: 21651511; http://dx.doi.org/10.1111/j.1399-0004.2011.01723.x - DOI - PMC - PubMed
-
- Jacquemont S, Hagerman RJ, Leehey MA, Hall DA, Levine RA, Brunberg JA, Zhang L, Jardini T, Gane LW, Harris SW, et al. Penetrance of the fragile x-associated tremor/ataxia syndrome in a premutation carrier population. JAMA 2004; 291:460–9; PMID: 14747503; http://dx.doi.org/10.1001/jama.291.4.460 - DOI - PubMed
-
- Hunsaker MR, Greco CM, Spath MA, Smits AP, Navarro CS, Tassone F, Kros JM, Severijnen LA, Berry-Kravis EM, Berman RF, et al. Widespread non-central nervous system organ pathology in fragile X premutation carriers with fragile X-associated tremor/ataxia syndrome and CGG knock-in mice. ActaNeuropathol 2011; 122:467–79; PMID: 21785977; http://dx.doi.org/10.1007/s00401-011-0860-9 - DOI - PMC - PubMed
-
- Hagerman R, Hagerman P. Advances in clinical and molecular understanding of the FMR1 premutation and fragile X-associated tremor/ataxia syndrome. LancetNeurol 2013; 12:786–98; PMID: 23867198; http://dx.doi.org/10.1016/S1474-4422(13)70125-X - DOI - PMC - PubMed
-
- Tassone F, Long KP, Tong TH, Lo J, Gane LW, Berry-Kravis E, Nguyen D, Mu LY, Laffin J, Bailey DB Jr., et al. FMR1 CGG allele size and prevalence ascertained through newborn screening in the United States. GenomeMed 2012; 4:100; PMID: 23259642; http://dx.doi.org/10.1186/gm401 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous