

Lack of centrioles and primary cilia in STIL(-/-) mouse embryos
- PMID: 25486474
- PMCID: PMC4615128
- DOI: 10.4161/15384101.2014.946830
Lack of centrioles and primary cilia in STIL(-/-) mouse embryos
Abstract
Although most animal cells contain centrosomes, consisting of a pair of centrioles, their precise contribution to cell division and embryonic development is unclear. Genetic ablation of STIL, an essential component of the centriole replication machinery in mammalian cells, causes embryonic lethality in mice around mid gestation associated with defective Hedgehog signaling. Here, we describe, by focused ion beam scanning electron microscopy, that STIL(-/-) mouse embryos do not contain centrioles or primary cilia, suggesting that these organelles are not essential for mammalian development until mid gestation. We further show that the lack of primary cilia explains the absence of Hedgehog signaling in STIL(-/-) cells. Exogenous re-expression of STIL or STIL microcephaly mutants compatible with human survival, induced non-templated, de novo generation of centrioles in STIL(-/-) cells. Thus, while the abscence of centrioles is compatible with mammalian gastrulation, lack of centrioles and primary cilia impairs Hedgehog signaling and further embryonic development.
Keywords: CDK6, cyclin-dependent kinase 6; CEP, centrosomal protein; COILEDX, coiled-coil domain deletion; E, embryonic day; FIB/SEM, focused ion beam scanning electron microscopy; MCPH, autosomal recessive primary microcephaly; MEFs, mouse embryonic fibroblasts; MTOC, microtubule organizing center; PLK4, polo kinase 4; SHH, sonic hedgehog; STAN, STIL/ANA2; STANX, STAN domain deletion; STIL; STIL, SCL/TAL1 interrupting locus; centriole; centrosome; electron microscopy; embryo; microcephaly; nm, nanometer; siRNA, small interfering RNA.
Figures
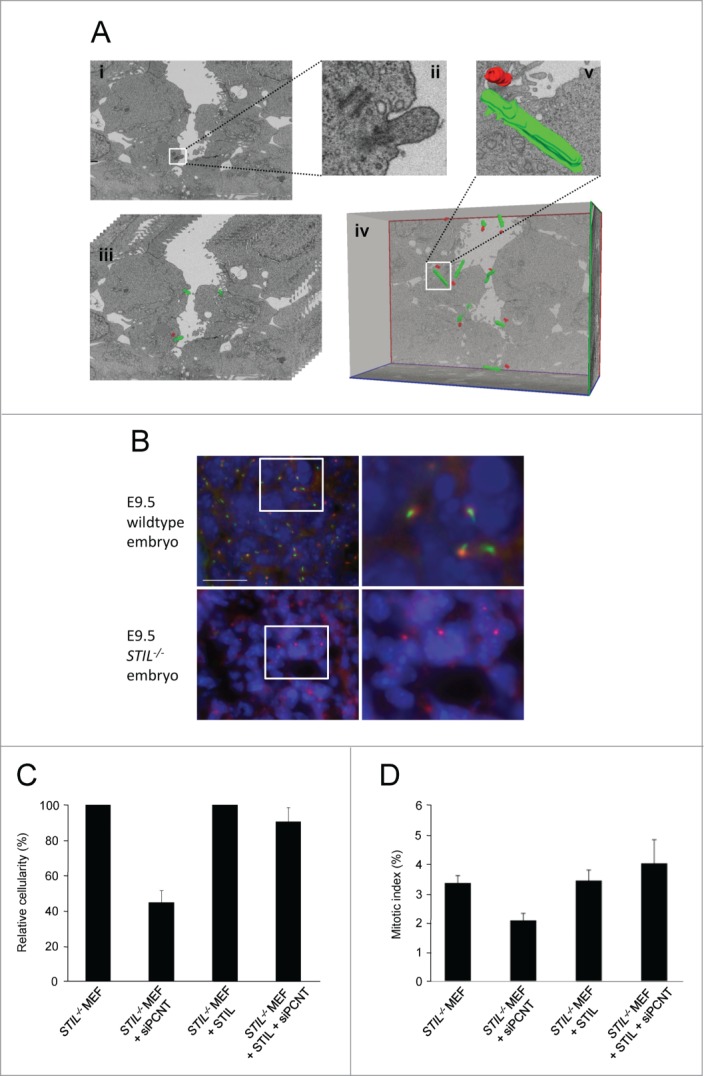
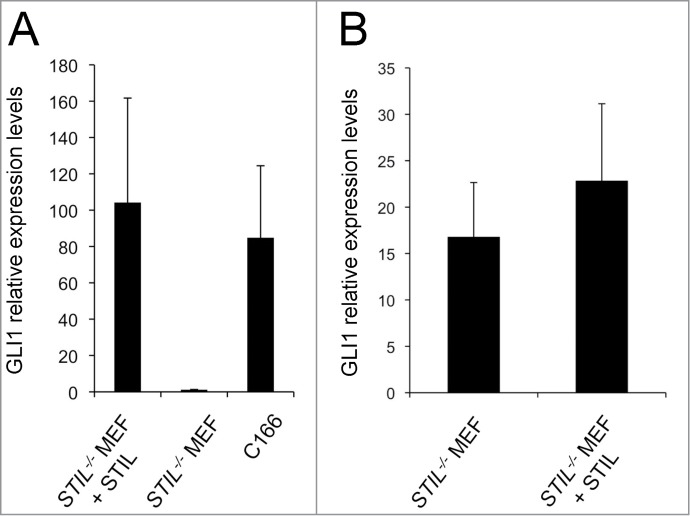
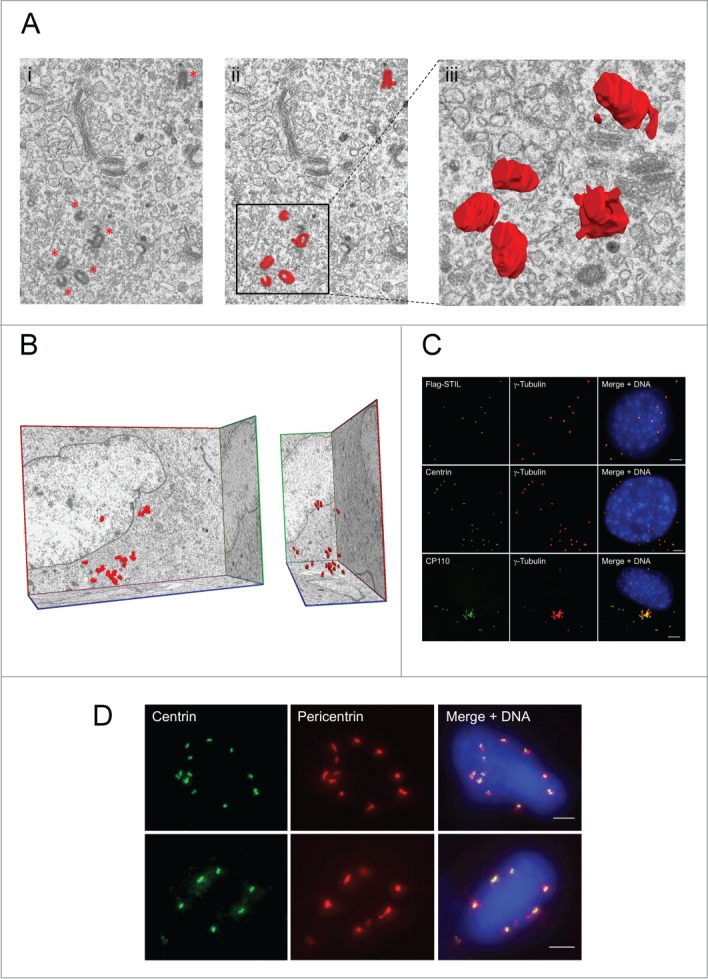
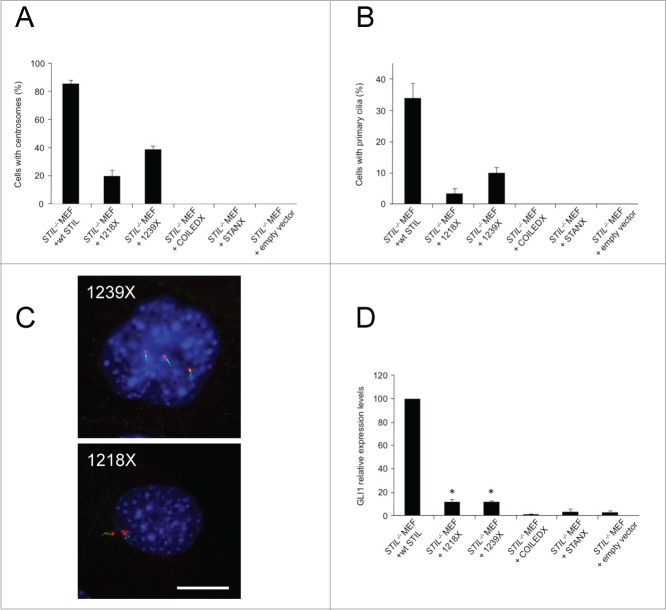
Comment in
-
Centrioles in the mouse: cilia and beyond.Cell Cycle. 2014;13(18):2809. doi: 10.4161/15384101.2014.954450. Cell Cycle. 2014. PMID: 25486466 Free PMC article. No abstract available.
References
-
- Azimzadeh J, Marshall WF. Building the centriole. Curr Biol 2010; 20:R816-25; PMID:20869612; http://dx.doi.org/ 10.1016/j.cub.2010.08.010 - DOI - PMC - PubMed
-
- Nigg EA, Stearns T. The centrosome cycle: centriole biogenesis, duplication and inherent asymmetries. Nat Cell Biol 2011; 13:1154-60; PMID:21968988; http://dx.doi.org/ 10.1038/ncb2345 - DOI - PMC - PubMed
-
- Dammermann A, Müller-Reichert T, Pelletier L, Habermann B, Desai A, Oegema K. Centriole assembly requires both centriolar and pericentriolar material proteins. Dev Cell 2004; 7:815-29; PMID:15572125; http://dx.doi.org/ 10.1016/j.devcel.2004.10.015 - DOI - PubMed
-
- Delattre M, Leidel S, Wani K, Baumer K, Bamat J, Schnabel H, Feichtinger R, Schnabel R, Gönczy P. Centriolar SAS-5 is required for centrosome duplication in C. elegans. Nat Cell Biol 2004; 6:656-64; PMID:15232593; http://dx.doi.org/ 10.1038/ncb1146 - DOI - PubMed
-
- Leidel S, Gönczy P. SAS-4 is essential for centrosome duplication in C elegans and is recruited to daughter centrioles once per cell cycle. Dev Cell 2003; 4:431-9; PMID:12636923; http://dx.doi.org/ 10.1016/S1534-5807(03)00062-5 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous