

Phosphorylation of ribosomal protein S6 kinase 1 at Thr421/Ser424 and dephosphorylation at Thr389 regulates SP600125-induced polyploidization of megakaryocytic cell lines
- PMID: 25486532
- PMCID: PMC4259319
- DOI: 10.1371/journal.pone.0114389
Phosphorylation of ribosomal protein S6 kinase 1 at Thr421/Ser424 and dephosphorylation at Thr389 regulates SP600125-induced polyploidization of megakaryocytic cell lines
Abstract
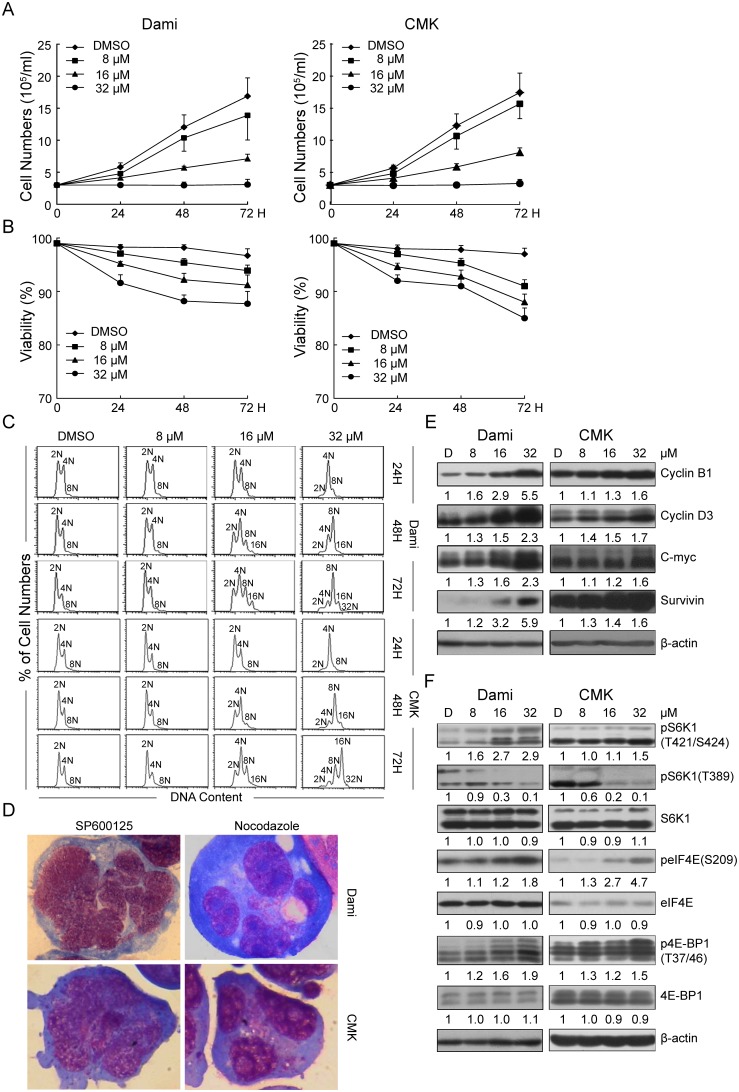
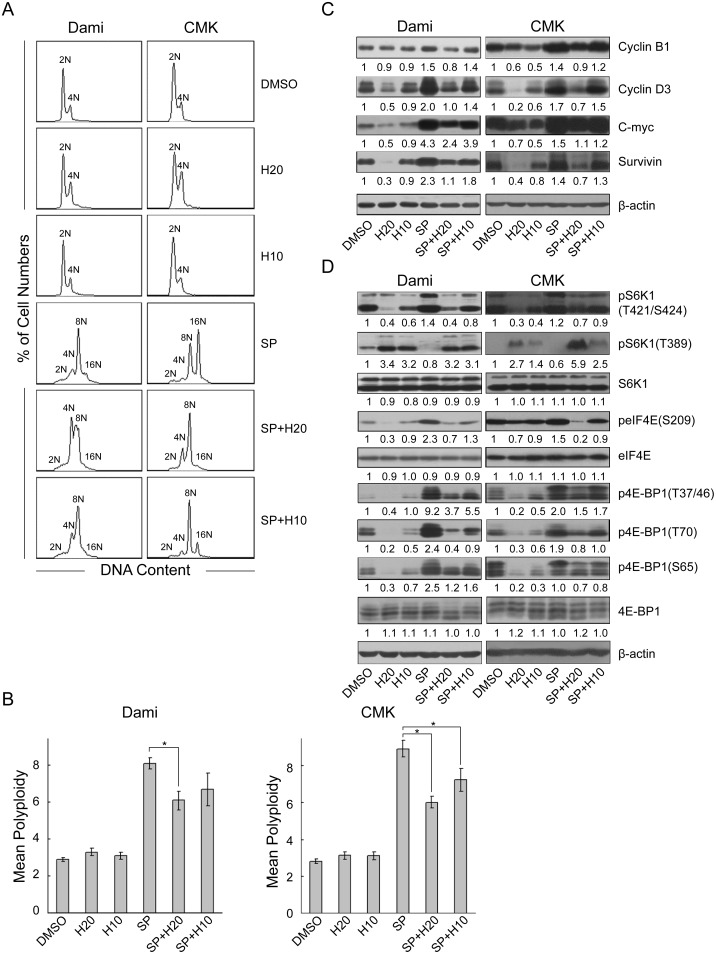
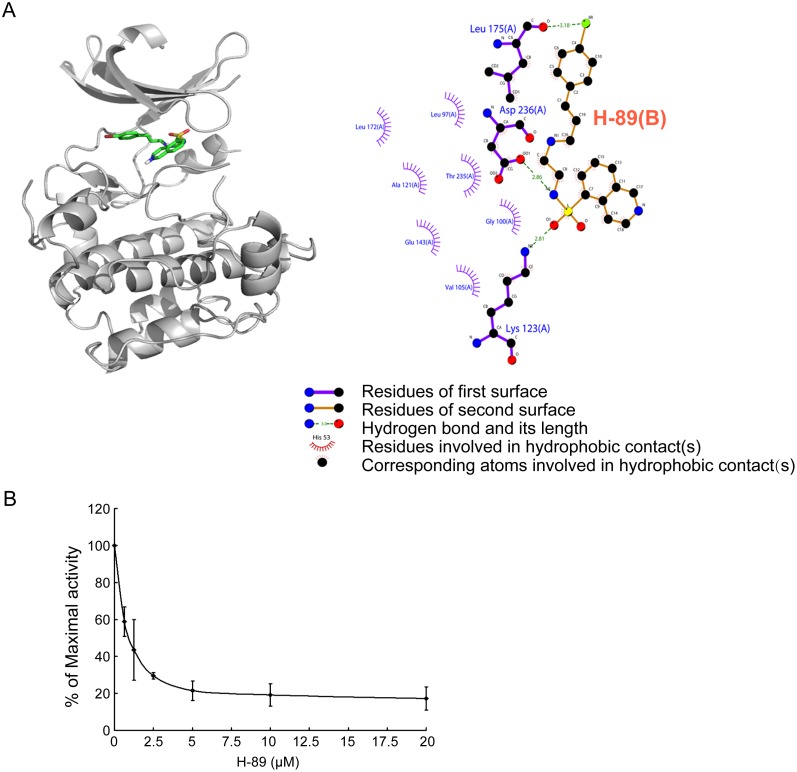
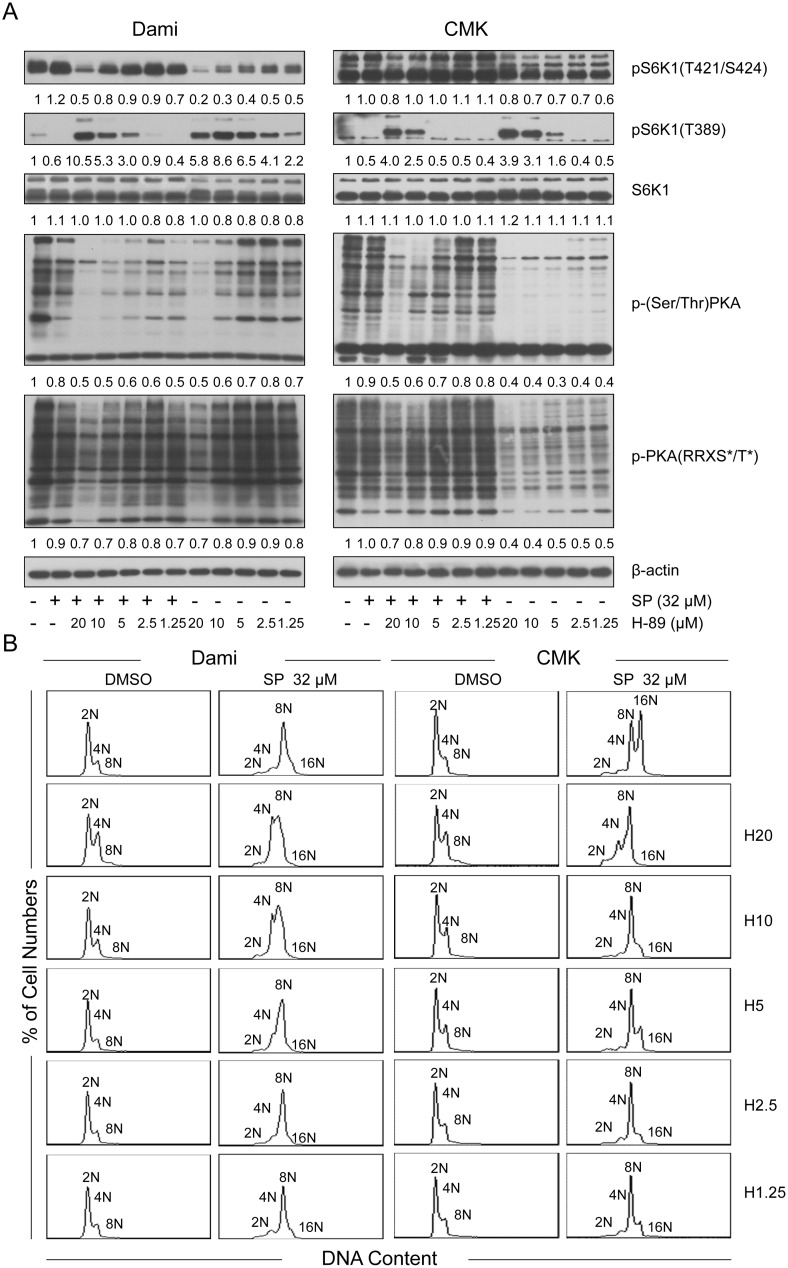
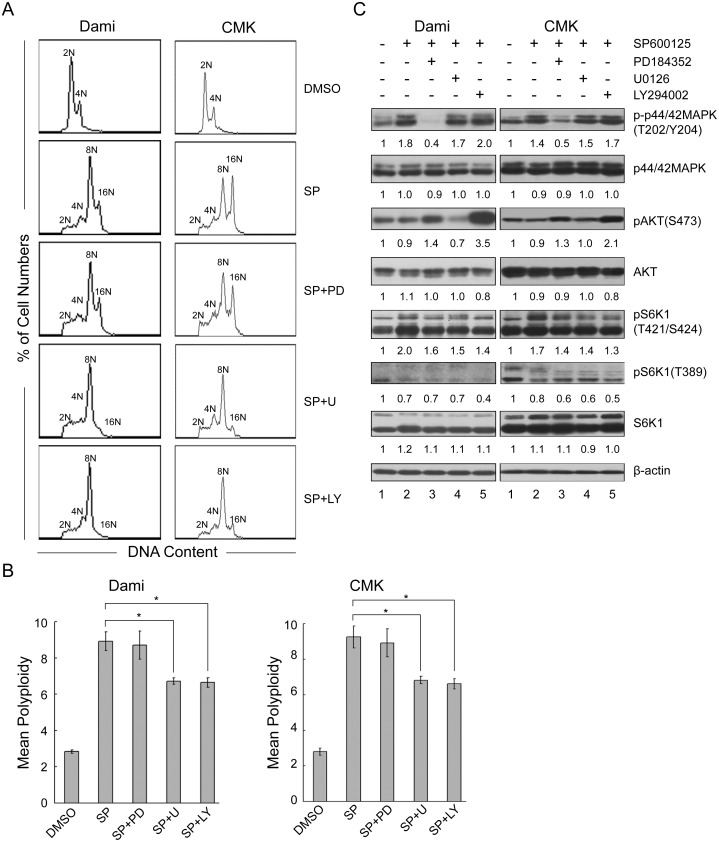
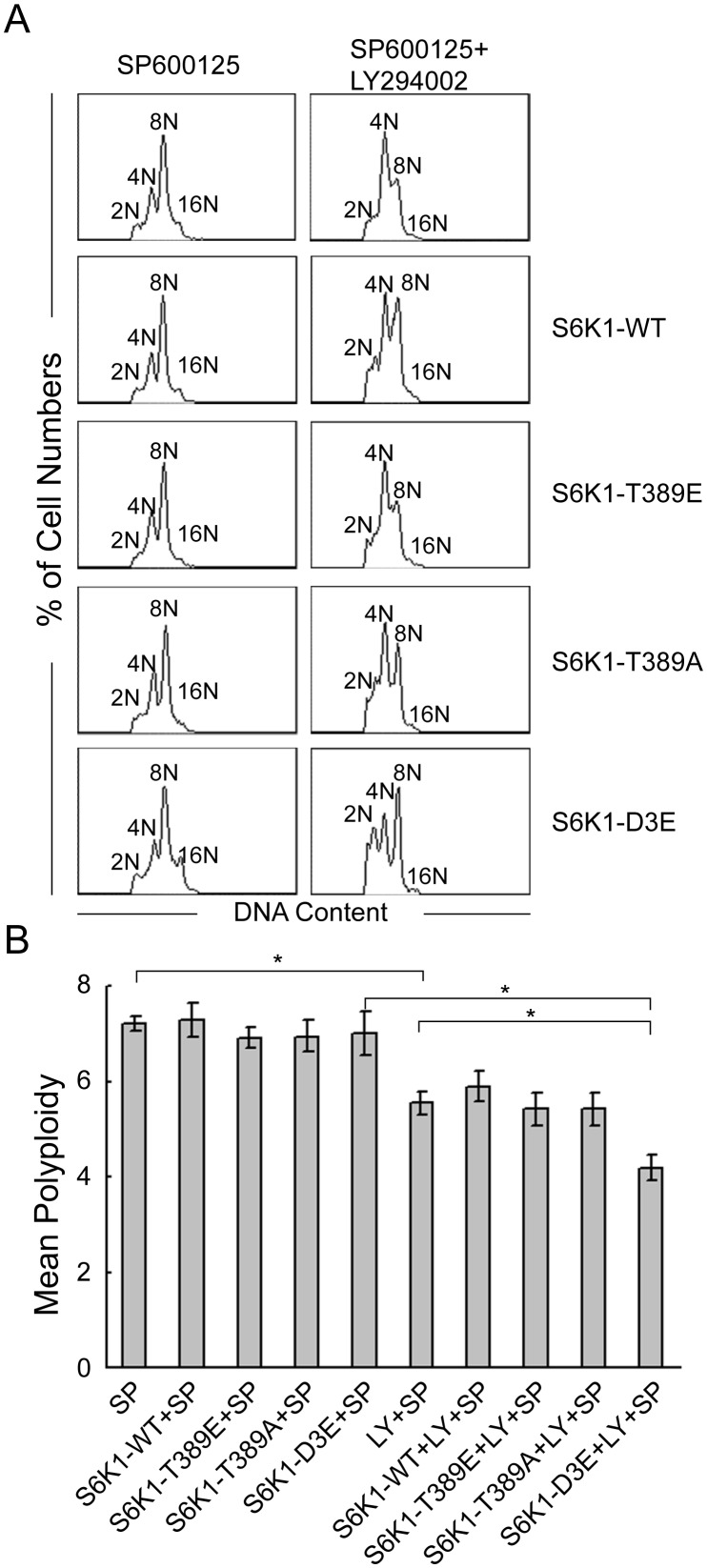
Megakaryocytes (MKs) are one of the few cell types that become polyploid; however, the mechanisms by which these cells are designated to become polyploid are not fully understood. In this investigation, we successfully established two relatively synchronous polyploid cell models by inducing Dami and CMK cells with SP600125. We found that SP600125 induced the polyploidization of Dami and CMK cells, concomitant with the phosphorylation of ribosomal protein S6 kinase 1 (S6K1) at Thr421/Ser424 and dephosphorylation at Thr389. The polyploidization was partially blocked by H-89, a cAMP-dependent protein kinase (PKA) inhibitor, through direct binding to S6K1, leading to dephosphorylation at Thr421/Ser424 and phosphorylation at Thr389, independent of PKA. Overexpression of a rapamycin-resistant mutant of S6K1 further enhanced the inhibitory effect of LY294002 on the SP600125-induced polyploidization of Dami and CMK cells. SP600125 also induced the polyploidization of Meg-01 cells, which are derived from a patient with chronic myelogenous leukemia, without causing a significant change in S6K1 phosphorylation. Additionally, SP600125 induced the polyploidization of HEL cells, which are derived from a patient with erythroleukemia, and phosphorylation at Thr389 of S6K1 was detected. However, the polyploidization of both Meg-01 cells and HEL cells as a result of SP600125 treatment was lower than that of SP600125-induced Dami and CMK cells, and it was not blocked by H-89 despite the increased phosphorylation of S6K1 at Thr389 in both cell lines in response to H-89. Given that the Dami and CMK cell lines were derived from patients with acute megakaryocytic leukemia (AMKL) and expressed high levels of platelet-specific antigens, our data suggested that SP600125-induced polyploidization is cell-type specific, that these cell lines were more differentiated, and that phosphorylation at Thr421/Ser424 and dephosphorylation at Thr389 of S6K1 may play an important role in the SP600125-induced polyploidization of these cell lines synergistically with other signaling pathways.
Conflict of interest statement
Figures
References
-
- Bluteau D, Lordier L, Di Stefano A, Chang Y, Raslova H, et al. (2009) Regulation of megakaryocyte maturation and platelet formation. J Thromb Haemost 7 Suppl 1: 227–234. - PubMed
-
- Vitrat N, Cohen-Solal K, Pique C, Le Couedic JP, Norol F, et al. (1998) Endomitosis of human megakaryocytes are due to abortive mitosis. Blood 91:3711–3723. - PubMed
-
- Asnaghi L, Bruno P, Priulla M, Nicolin A (2004) mTOR: a protein kinase switching between life and death. Pharmacol Res 50:545–549. - PubMed
-
- Guerriero R, Parolini I, Testa U, Samoggia P, Petrucci E, et al. (2006) Inhibition of TPO-induced MEK or mTOR activity induces opposite effects on the ploidy of human differentiating megakaryocytes. J Cell Sci 119:744–752. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
