

Cyclin B3 and dynein heavy chain cooperate to increase fitness in the absence of mdf-1/MAD1 in Caenorhabditis elegans
- PMID: 25486568
- PMCID: PMC4614836
- DOI: 10.4161/15384101.2014.949491
Cyclin B3 and dynein heavy chain cooperate to increase fitness in the absence of mdf-1/MAD1 in Caenorhabditis elegans
Abstract
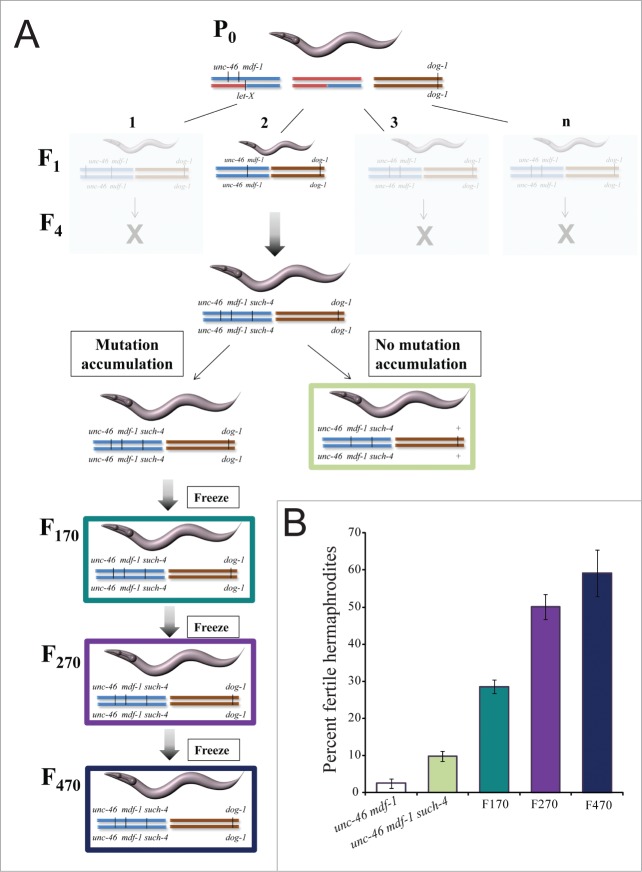
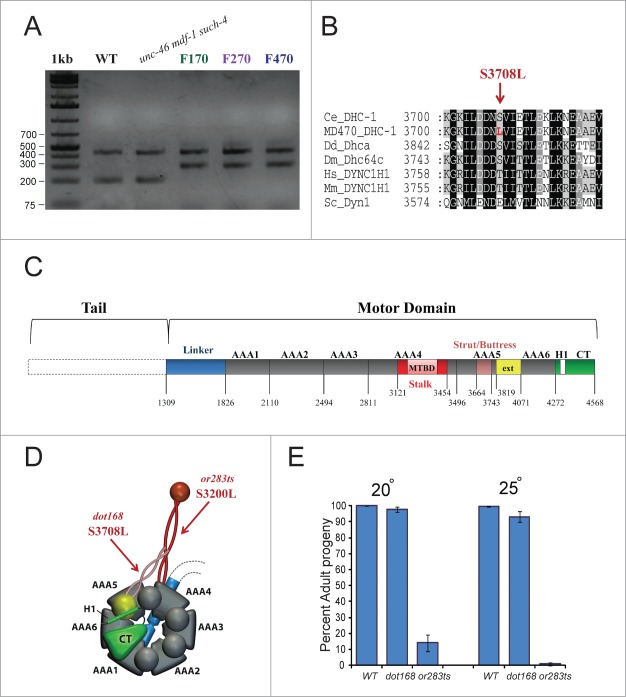
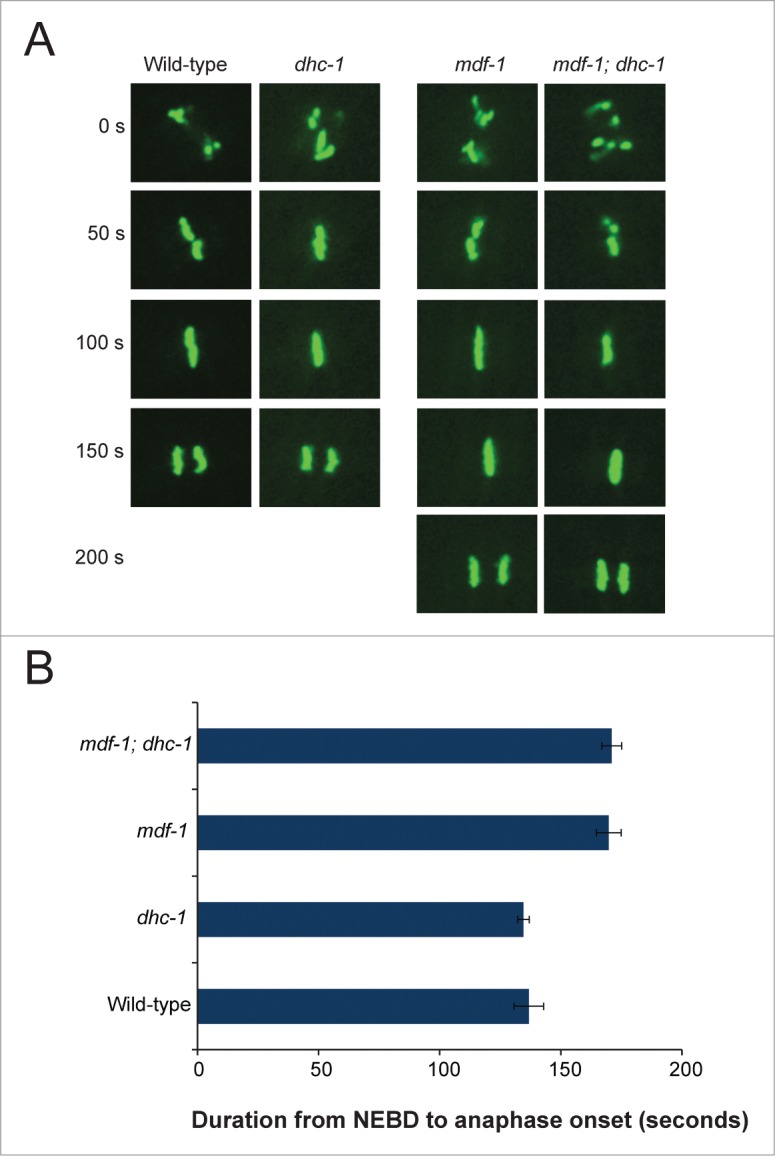
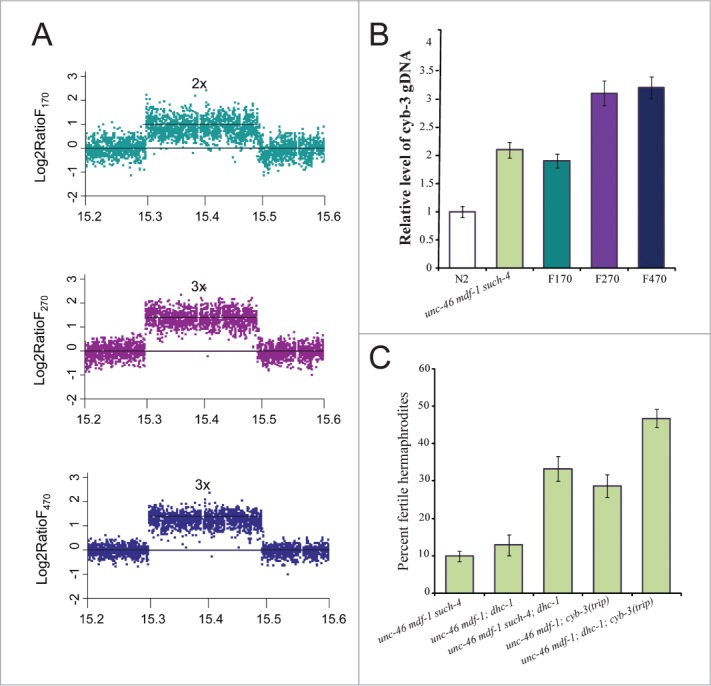
Spindle assembly checkpoint (SAC) ensures genome stability by delaying anaphase onset until all the chromosomes have achieved proper spindle attachment. Once correct attachment has been achieved, SAC must be silenced. In the absence of mdf-1/MAD1, an essential SAC component, Caenorhabditis elegans cannot propagate beyond 3 generations. Previously, in a dog-1(gk10)/FANCJ mutator background, we isolated a suppressor of mdf-1(gk2) sterility (such-4) which allowed indefinite propagation in the absence of MDF-1. We showed that such-4 is a Cyclin B3 (cyb-3) duplication. Here we analyze mdf-1 such-4; dog-1, which we propagated for 470 generations, with freezing of samples for long time storage at F170 and F270. Phenotypic analysis of this strain revealed additional suppression of sterility in the absence of MDF-1, beyond the effects of such-4. We applied oligonucleotide array Comparative Genomic Hybridization (oaCGH) and whole genome sequencing (WGS) and identified a further amplification of cyb-3 (triplication) and a new missense mutation in dynein heavy chain (dhc-1). We show that dhc-1(dot168) suppresses the mdf-1(gk2), and is the second cloned suppressor, next to cyb-3 duplication, that does not cause a delay in anaphase onset. We also show that amplification of cyb-3 and dhc-1(dot168) cooperate to increase fitness in the absence of MDF-1.
Keywords: APC/C, anaphase promoting complex/cyclosome; CIN, chromosome instability; Cyclin B3 (cyb-3); EMS, ethyl methanesulfonate; Him, high incidence of males; SAC, spindle assembly checkpoint; WGS, whole genome sequencing; dynein heavy chain (dhc-1); genome stability; mdf-1/MAD1; oaCGH, oligonucleotide array Comparative Genomic Hybridization; spindle assembly checkpoint (SAC).
Figures
Comment in
-
ATM deficiency: revealing the pathways to cancer.Cell Cycle. 2014;13(19):2992. doi: 10.4161/15384101.2014.959849. Cell Cycle. 2014. PMID: 25486558 Free PMC article. No abstract available.
References
-
- Musacchio A, Salmon ED. The spindle-assembly checkpoint in space and time. Nat Rev Mol Cell Biol 2007; 8:379-93; PMID:17426725; http://dx.doi.org/ 10.1038/nrm2163 - DOI - PubMed
-
- Hauf S. The spindle assembly checkpoint: progress and persistent puzzles. Biochem Soc Trans 2013; 41:1755-60; PMID:24256287 - PubMed
-
- Kitagawa R, Rose AM. Components of the spindle-assembly checkpoint are essential in Caenorhabditis elegans. Nat Cell Biol 1999; 1:514-21; PMID:10587648; http://dx.doi.org/ 10.1038/70309 - DOI - PubMed
-
- Oegema K, Desai A, Rybina S, Kirkham M, Hyman AA. Functional analysis of kinetochore assembly in Caenorhabditis elegans. J Cell Biol 2001; 153:1209-26; PMID:11402065; http://dx.doi.org/ 10.1083/jcb.153.6.1209 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous