

Targeted deletion of the ara operon of Salmonella typhimurium enhances L-arabinose accumulation and drives PBAD-promoted expression of anti-cancer toxins and imaging agents
- PMID: 25486570
- PMCID: PMC4613543
- DOI: 10.4161/15384101.2014.949527
Targeted deletion of the ara operon of Salmonella typhimurium enhances L-arabinose accumulation and drives PBAD-promoted expression of anti-cancer toxins and imaging agents
Abstract
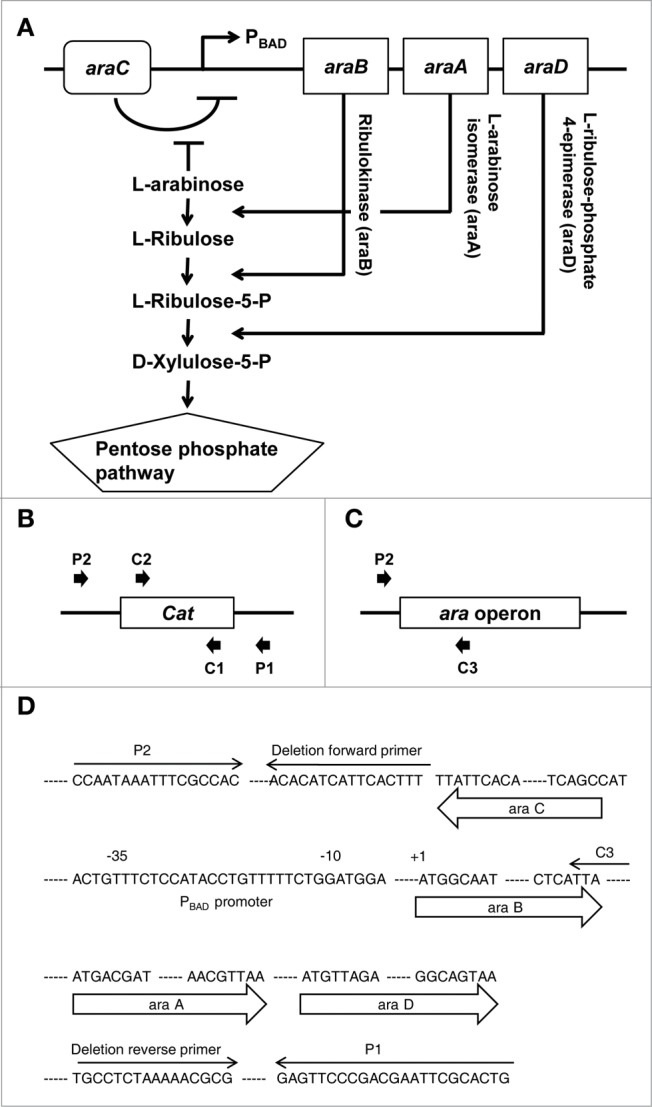
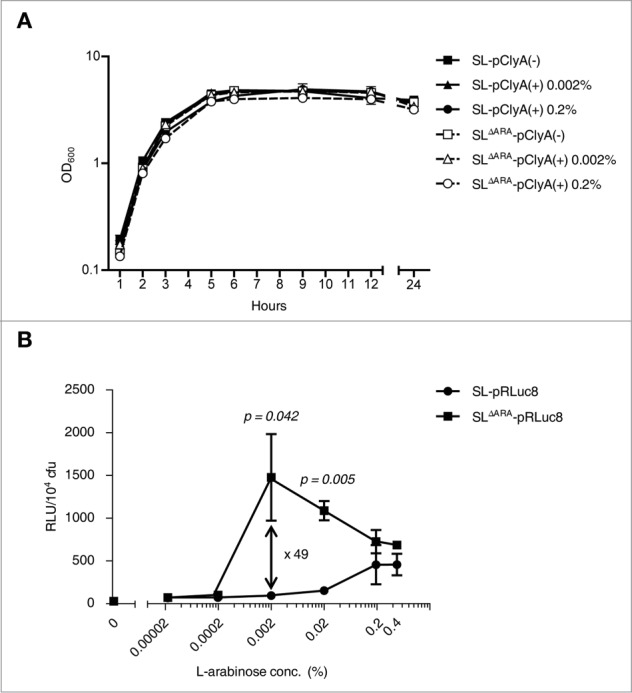
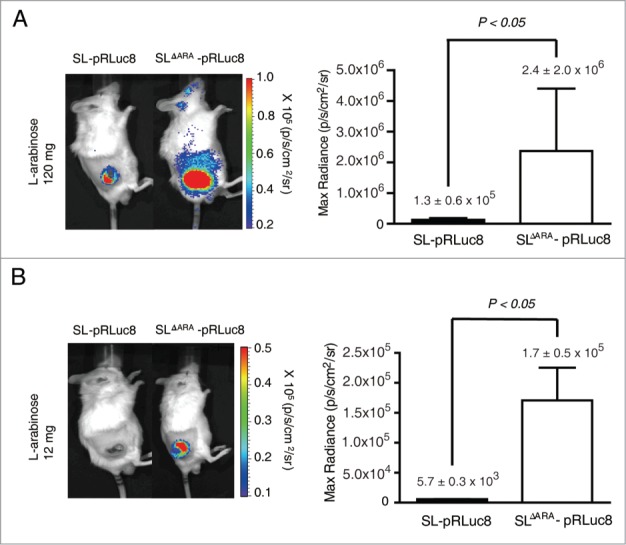
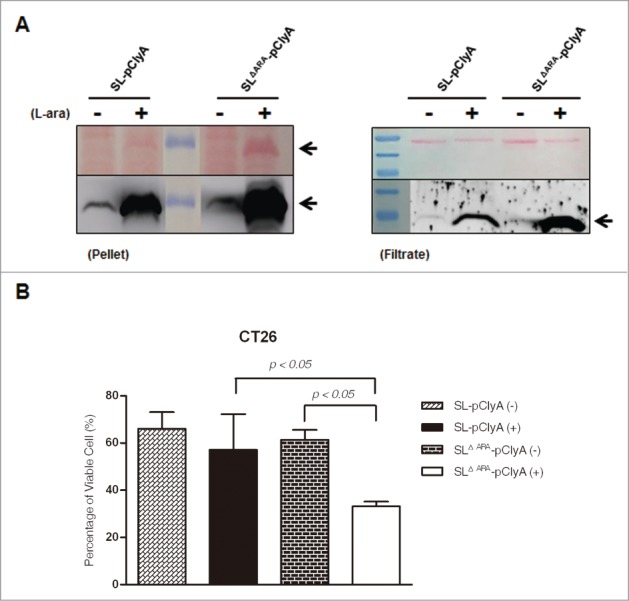
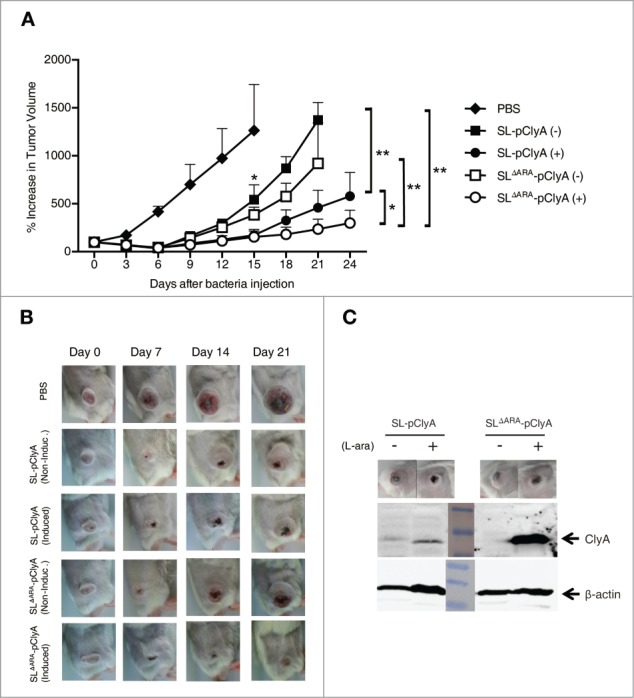
Tumor-specific expression of antitumor drugs can be achieved using attenuated Salmonella typhimurium harboring the PBAD promoter, which is induced by L-arabinose. However, L-arabinose does not accumulate because it is metabolized to D-xylulose-5-P by enzymes encoded by the ara operon in Salmonellae. To address this problem, we developed an engineered strain of S. typhimurium in which the ara operon is deleted. Linear DNA transformation was performed using λ red recombinase to exchange the ara operon with linear DNA carrying an antibiotic-resistance gene with homology to regions adjacent to the ara operon. The ara operon-deleted strain and its parental strain were transformed with a plasmid encoding Renilla luciferase variant 8 (RLuc8) or cytolysin A (clyA) under the control of the PBAD promoter. Luciferase assays demonstrated that RLuc8 expression was 49-fold higher in the ara operon-deleted S. typhimurium than in the parental strain after the addition of L-arabinose. In vivo bioluminescence imaging showed that the tumor tissue targeted by the ara operon-deleted Salmonella had a stronger imaging signal (~30-fold) than that targeted by the parental strain. Mice with murine colon cancer (CT26) that had been injected with the ara operon-deleted S. typhimurium expressing clyA showed significant tumor suppression. The present report demonstrates that deletion of the ara operon of S. typhimurium enhances L-arabinose accumulation and thereby drives PBAD-promoted expression of cytotoxic agents and imaging agents. This is a promising approach for tumor therapy and imaging.
Keywords: L-arabinose; PBAD promoter; Salmonella typhimurium; ara operon; bacterial cancer therapy; bioluminescence; cytolysin A.
Figures
Comment in
-
Tuning Salmonella for cancer therapy.Cell Cycle. 2015;14(8):1138. doi: 10.1080/15384101.2015.1006561. Cell Cycle. 2015. PMID: 25590787 Free PMC article. No abstract available.
Similar articles
-
Remote control of tumour-targeted Salmonella enterica serovar Typhimurium by the use of L-arabinose as inducer of bacterial gene expression in vivo.Cell Microbiol. 2007 Jun;9(6):1529-37. doi: 10.1111/j.1462-5822.2007.00890.x. Epub 2007 Feb 9. Cell Microbiol. 2007. PMID: 17298393
-
Radiotherapy combined with an engineered Salmonella typhimurium inhibits tumor growth in a mouse model of colon cancer.Exp Anim. 2016 Nov 1;65(4):413-418. doi: 10.1538/expanim.16-0033. Epub 2016 Jun 10. Exp Anim. 2016. PMID: 27301721 Free PMC article.
-
The Salmonella typhimurium mar locus: molecular and genetic analyses and assessment of its role in virulence.J Bacteriol. 1997 Mar;179(6):1857-66. doi: 10.1128/jb.179.6.1857-1866.1997. J Bacteriol. 1997. PMID: 9068629 Free PMC article.
-
Factors modulating expression of Renilla luciferase from control plasmids used in luciferase reporter gene assays.Anal Biochem. 2010 Jan 15;396(2):167-72. doi: 10.1016/j.ab.2009.09.043. Epub 2009 Sep 27. Anal Biochem. 2010. PMID: 19788887 Free PMC article. Review.
-
Metabolism of L-arabinose in plants.J Plant Res. 2016 Sep;129(5):781-792. doi: 10.1007/s10265-016-0834-z. Epub 2016 May 24. J Plant Res. 2016. PMID: 27220955 Free PMC article. Review.
Cited by
-
Salmonella enterica and outer membrane vesicles are current and future options for cancer treatment.Front Cell Infect Microbiol. 2023 Dec 5;13:1293351. doi: 10.3389/fcimb.2023.1293351. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 38116133 Free PMC article. Review.
-
Attenuated mutant strain of Salmonella Typhimurium lacking the ZnuABC transporter contrasts tumor growth promoting anti-cancer immune response.Oncotarget. 2015 Jul 10;6(19):17648-60. doi: 10.18632/oncotarget.3893. Oncotarget. 2015. PMID: 26158862 Free PMC article.
-
A versatile regulatory toolkit of arabinose-inducible artificial transcription factors for Enterobacteriaceae.Commun Biol. 2023 Oct 3;6(1):1005. doi: 10.1038/s42003-023-05363-3. Commun Biol. 2023. PMID: 37789111 Free PMC article.
-
Recent advances in bacterial therapeutics based on sense and response.Acta Pharm Sin B. 2023 Mar;13(3):1014-1027. doi: 10.1016/j.apsb.2022.09.015. Epub 2022 Sep 25. Acta Pharm Sin B. 2023. PMID: 36970195 Free PMC article. Review.
-
Salmonella-Mediated Cancer Therapy: Roles and Potential.Nucl Med Mol Imaging. 2017 Jun;51(2):118-126. doi: 10.1007/s13139-016-0415-z. Epub 2016 Jun 1. Nucl Med Mol Imaging. 2017. PMID: 28559936 Free PMC article. Review.
References
-
- Kishimoto H, Aki R, Urata Y, Bouvet M, Momiyama M, Tanaka N, Fujiwara T, Hoffman RM. Tumor-selective, adenoviral-mediated GFP genetic labeling of human cancer in the live mouse reports future recurrence after resection. Cell Cycle 2011; 10:2737-41; PMID:21785265; http://dx.doi.org/10.4161/cc.10.16.16756 - DOI - PMC - PubMed
-
- Zhao M, Yang M, Li XM, Jiang P, Baranov E, Li S, Xu M, Penman S, Hoffman RM. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc Natl Acad Sci U S A 2005; 102:755-60; PMID:15644448; http://dx.doi.org/10.1073/pnas.0408422102 - DOI - PMC - PubMed
-
- Yu YA, Shabahang S, Timiryasova TM, Zhang Q, Beltz R, Gentschev I, Goebel W, Szalay AA. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light-emitting proteins. Nat Biotechnol 2004; 22(3):313-20; PMID:14990953; http://dx.doi.org/10.1038/nbt937 - DOI - PubMed
-
- Pawelek JM, Low KB, Bermudes D. Bacteria as tumour-targeting vectors. Lancet Oncol 2003; 4(11):658; http://dx.doi.org/10.1016/S1470-2045(03)01194-X - DOI - PubMed
-
- Min JJ, Nguyen VH, Kim HJ, Hong Y, Choy HE. Quantitative bioluminescence imaging of tumor-targeting bacteria in living animals. Nat Protoc 2008; 3(4):629-36; PMID:18388945; http://dx.doi.org/10.1038/nprot.2008.32 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources