

The C-terminal 50 amino acid residues of dengue NS3 protein are important for NS3-NS5 interaction and viral replication
- PMID: 25488659
- PMCID: PMC4303688
- DOI: 10.1074/jbc.M114.607341
The C-terminal 50 amino acid residues of dengue NS3 protein are important for NS3-NS5 interaction and viral replication
Abstract
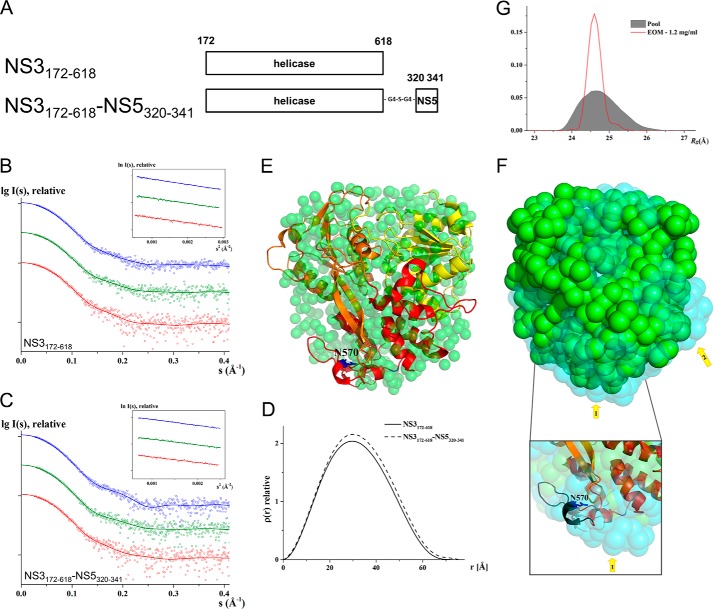
Dengue virus multifunctional proteins NS3 protease/helicase and NS5 methyltransferase/RNA-dependent RNA polymerase form part of the viral replication complex and are involved in viral RNA genome synthesis, methylation of the 5'-cap of viral genome, and polyprotein processing among other activities. Previous studies have shown that NS5 residue Lys-330 is required for interaction between NS3 and NS5. Here, we show by competitive NS3-NS5 interaction ELISA that the NS3 peptide spanning residues 566-585 disrupts NS3-NS5 interaction but not the null-peptide bearing the N570A mutation. Small angle x-ray scattering study on NS3(172-618) helicase and covalently linked NS3(172-618)-NS5(320-341) reveals a rigid and compact formation of the latter, indicating that peptide NS5(320-341) engages in specific and discrete interaction with NS3. Significantly, NS3:Asn-570 to alanine mutation introduced into an infectious DENV2 cDNA clone did not yield detectable virus by plaque assay even though intracellular double-stranded RNA was detected by immunofluorescence. Detection of increased negative-strand RNA synthesis by real time RT-PCR for the NS3:N570A mutant suggests that NS3-NS5 interaction plays an important role in the balanced synthesis of positive- and negative-strand RNA for robust viral replication. Dengue virus infection has become a global concern, and the lack of safe vaccines or antiviral treatments urgently needs to be addressed. NS3 and NS5 are highly conserved among the four serotypes, and the protein sequence around the pinpointed amino acids from the NS3 and NS5 regions are also conserved. The identification of the functionally essential interaction between the two proteins by biochemical and reverse genetics methods paves the way for rational drug design efforts to inhibit viral RNA synthesis.
Keywords: Dengue Virus; Flavivirus; Nonstructural Protein Interaction; Plus-stranded RNA Virus; Protein-Protein Interaction; Replication Complex; Viral Helicase; Viral Polymerase; Viral Protein; Viral Replication.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






Similar articles
-
Serotype-specific interactions among functional domains of dengue virus 2 nonstructural proteins (NS) 5 and NS3 are crucial for viral RNA replication.J Biol Chem. 2017 Jun 9;292(23):9465-9479. doi: 10.1074/jbc.M117.775643. Epub 2017 Apr 10. J Biol Chem. 2017. PMID: 28396347 Free PMC article.
-
The Transactions of NS3 and NS5 in Flaviviral RNA Replication.Adv Exp Med Biol. 2018;1062:147-163. doi: 10.1007/978-981-10-8727-1_11. Adv Exp Med Biol. 2018. PMID: 29845531 Review.
-
Substitution of NS5 N-terminal domain of dengue virus type 2 RNA with type 4 domain caused impaired replication and emergence of adaptive mutants with enhanced fitness.J Biol Chem. 2014 Aug 8;289(32):22385-400. doi: 10.1074/jbc.M114.584466. Epub 2014 Jun 5. J Biol Chem. 2014. PMID: 24904061 Free PMC article.
-
A Proline-Rich N-Terminal Region of the Dengue Virus NS3 Is Crucial for Infectious Particle Production.J Virol. 2016 May 12;90(11):5451-61. doi: 10.1128/JVI.00206-16. Print 2016 Jun 1. J Virol. 2016. PMID: 27009958 Free PMC article.
-
Towards the design of antiviral inhibitors against flaviviruses: the case for the multifunctional NS3 protein from Dengue virus as a target.Antiviral Res. 2008 Nov;80(2):94-101. doi: 10.1016/j.antiviral.2008.07.001. Epub 2008 Jul 30. Antiviral Res. 2008. PMID: 18674567 Review.
Cited by
-
Structural features of NS3 of Dengue virus serotypes 2 and 4 in solution and insight into RNA binding and the inhibitory role of quercetin.Acta Crystallogr D Struct Biol. 2017 May 1;73(Pt 5):402-419. doi: 10.1107/S2059798317003849. Epub 2017 Apr 19. Acta Crystallogr D Struct Biol. 2017. PMID: 28471365 Free PMC article.
-
Structure of the NS3 helicase from Zika virus.Nat Struct Mol Biol. 2016 Aug;23(8):752-4. doi: 10.1038/nsmb.3258. Epub 2016 Jul 11. Nat Struct Mol Biol. 2016. PMID: 27399257 Free PMC article.
-
Deciphering the interaction surface between the West Nile virus NS3 and NS5 proteins.Access Microbiol. 2024 Jun 26;6(6):000675.v3. doi: 10.1099/acmi.0.000675.v3. eCollection 2024. Access Microbiol. 2024. PMID: 39045235 Free PMC article.
-
Dual-Role Ubiquitination Regulation Shuttling the Entire Life Cycle of the Flaviviridae.Front Microbiol. 2022 May 6;13:835344. doi: 10.3389/fmicb.2022.835344. eCollection 2022. Front Microbiol. 2022. PMID: 35602051 Free PMC article. Review.
-
Potential Dual Role of West Nile Virus NS2B in Orchestrating NS3 Enzymatic Activity in Viral Replication.Viruses. 2021 Jan 31;13(2):216. doi: 10.3390/v13020216. Viruses. 2021. PMID: 33572517 Free PMC article.
References
-
- Bhatt S., Gething P. W., Brady O. J., Messina J. P., Farlow A. W., Moyes C. L., Drake J. M., Brownstein J. S., Hoen A. G., Sankoh O., Myers M. F., George D. B., Jaenisch T., Wint G. R., Simmons C. P., Scott T. W., Farrar J. J., Hay S. I. (2013) The global distribution and burden of dengue. Nature 496, 504–507 - PMC - PubMed
-
- Lindenbach B. D., Thiel H., Rice C. M. (2007) in Fields Virology (Knipe D. M., Howley P. M., eds) 5th Ed., pp. 1101–1152, Wolters Kluwer/Lippincott Williams & Wilkins, Philadelphia
-
- Bartholomeusz A. I., Wright P. J. (1993) Synthesis of dengue virus RNA in vitro: initiation and the involvement of proteins NS3 and NS5. Arch. Virol. 128, 111–121 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
