

BRCA2 diffuses as oligomeric clusters with RAD51 and changes mobility after DNA damage in live cells
- PMID: 25488918
- PMCID: PMC4259808
- DOI: 10.1083/jcb.201405014
BRCA2 diffuses as oligomeric clusters with RAD51 and changes mobility after DNA damage in live cells
Erratum in
-
BRCA2 diffuses as oligomeric clusters with RAD51 and changes mobility after DNA damage in live cells.J Cell Biol. 2015 Mar 16;208(6):857. doi: 10.1083/jcb.20140501402182015c. J Cell Biol. 2015. PMID: 25778924 Free PMC article. No abstract available.
Abstract
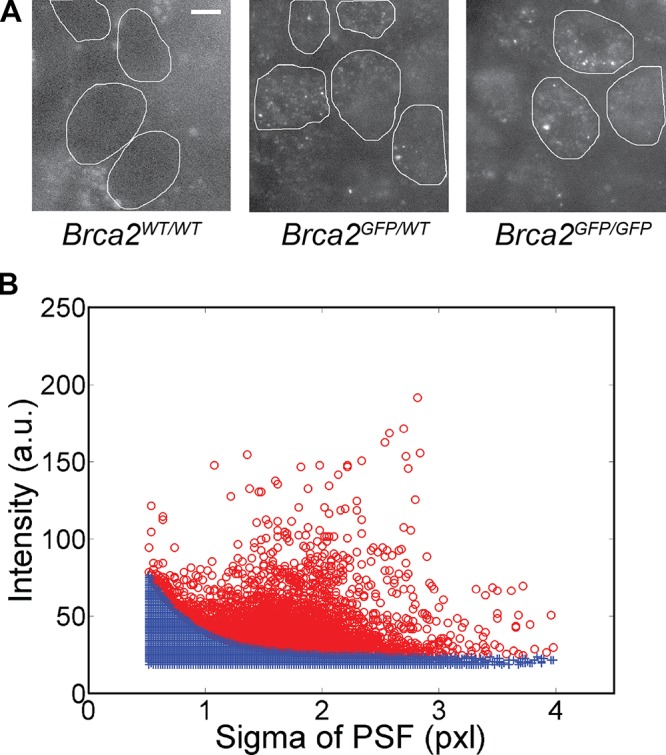
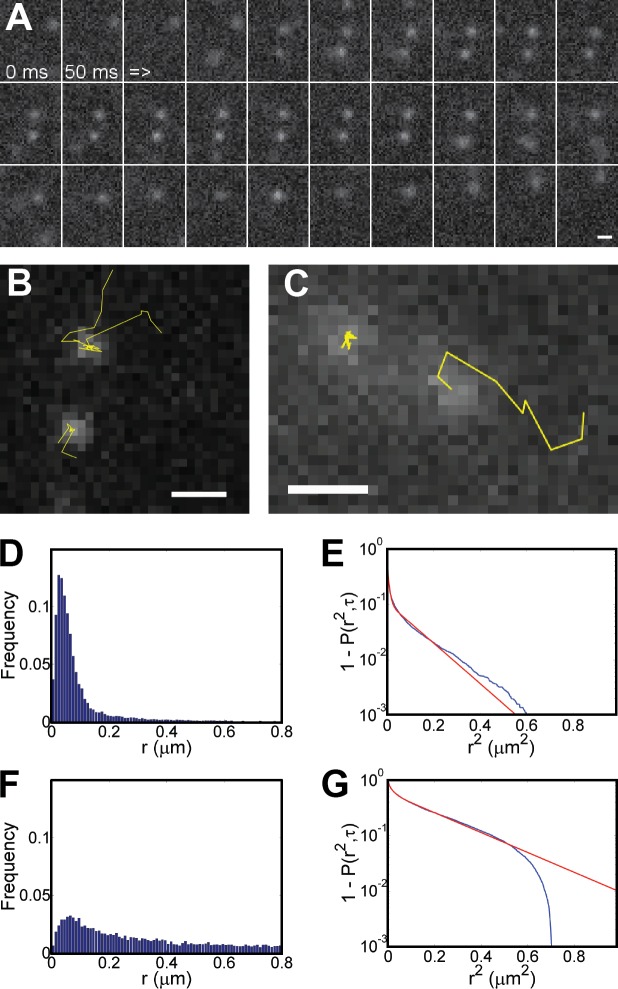
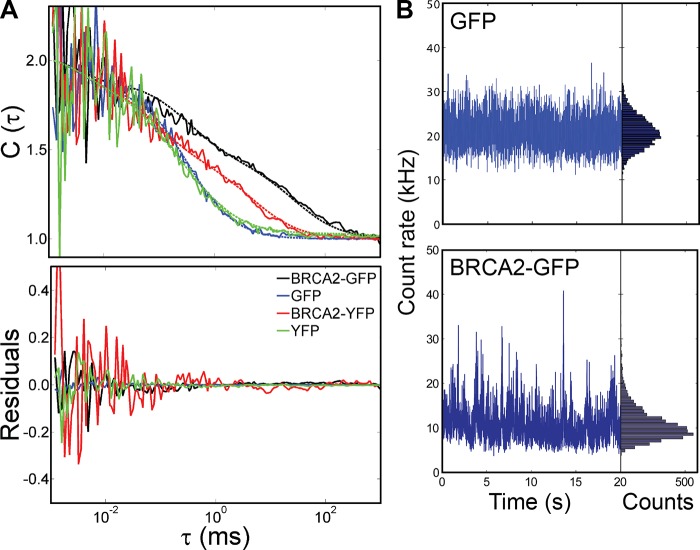
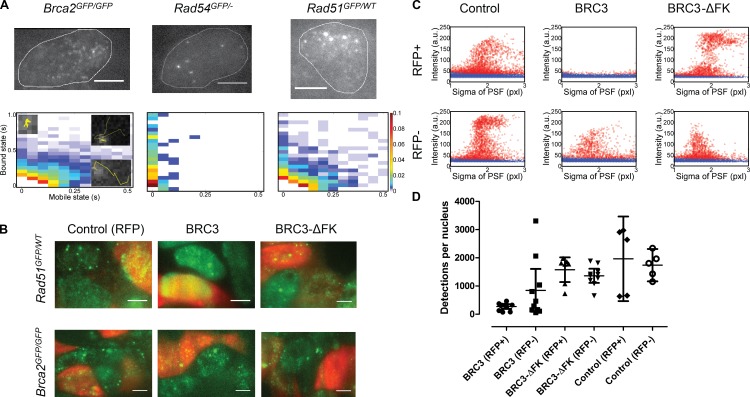
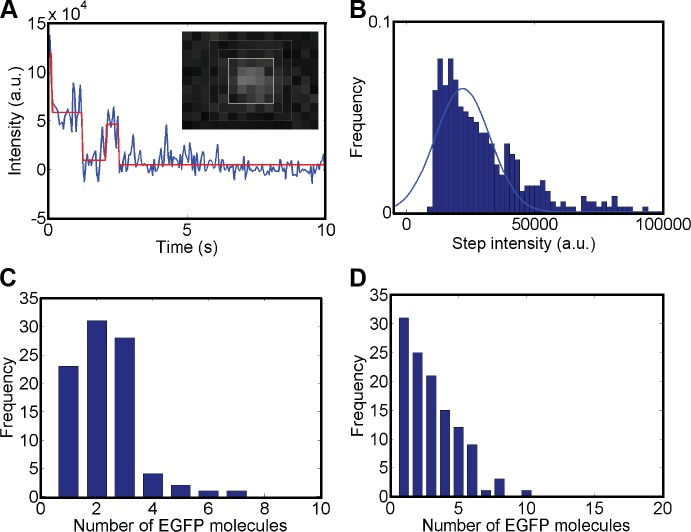
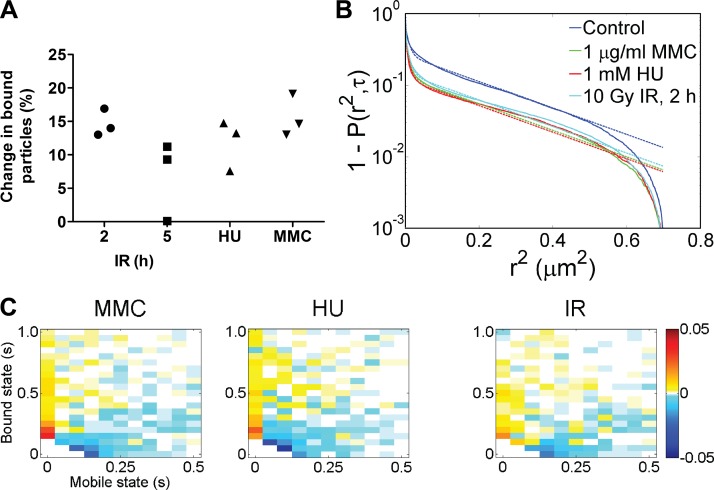
Genome maintenance by homologous recombination depends on coordinating many proteins in time and space to assemble at DNA break sites. To understand this process, we followed the mobility of BRCA2, a critical recombination mediator, in live cells at the single-molecule level using both single-particle tracking and fluorescence correlation spectroscopy. BRCA2-GFP and -YFP were compared to distinguish diffusion from fluorophore behavior. Diffusive behavior of fluorescent RAD51 and RAD54 was determined for comparison. All fluorescent proteins were expressed from endogenous loci. We found that nuclear BRCA2 existed in oligomeric clusters, and exhibited heterogeneous mobility. DNA damage increased BRCA2 transient binding, presumably including binding to damaged sites. Despite its very different size, RAD51 displayed mobility similar to BRCA2, which indicates physical interaction between these proteins both before and after induction of DNA damage. We propose that BRCA2-mediated sequestration of nuclear RAD51 serves to prevent inappropriate DNA interactions and that all RAD51 is delivered to DNA damage sites in association with BRCA2.
© 2014 Reuter et al.
Figures
Similar articles
-
Recovery of deficient homologous recombination in Brca2-depleted mouse cells by wild-type Rad51 expression.DNA Repair (Amst). 2009 Feb 1;8(2):170-81. doi: 10.1016/j.dnarep.2008.10.002. Epub 2008 Nov 18. DNA Repair (Amst). 2009. PMID: 18992372
-
Identification of Rad51 regulation by BRCA2 using Caenorhabditis elegans BRCA2 and bimolecular fluorescence complementation analysis.Biochem Biophys Res Commun. 2007 Nov 3;362(4):958-64. doi: 10.1016/j.bbrc.2007.08.083. Epub 2007 Aug 27. Biochem Biophys Res Commun. 2007. PMID: 17767921
-
DNA damage regulates the mobility of Brca2 within the nucleoplasm of living cells.Proc Natl Acad Sci U S A. 2010 Dec 14;107(50):21937-42. doi: 10.1073/pnas.1009577107. Epub 2010 Nov 22. Proc Natl Acad Sci U S A. 2010. PMID: 21098284 Free PMC article.
-
Suppression of the DNA repair defects of BRCA2-deficient cells with heterologous protein fusions.Proc Natl Acad Sci U S A. 2006 Jun 6;103(23):8768-73. doi: 10.1073/pnas.0600298103. Epub 2006 May 26. Proc Natl Acad Sci U S A. 2006. PMID: 16731627 Free PMC article.
-
BRCA2: a universal recombinase regulator.Oncogene. 2007 Dec 10;26(56):7720-30. doi: 10.1038/sj.onc.1210870. Oncogene. 2007. PMID: 18066084 Review.
Cited by
-
Repair of DNA double-strand breaks in plant meiosis: role of eukaryotic RecA recombinases and their modulators.Plant Reprod. 2023 Mar;36(1):17-41. doi: 10.1007/s00497-022-00443-6. Epub 2022 Jun 1. Plant Reprod. 2023. PMID: 35641832 Review.
-
The protein phosphatase EYA4 promotes homologous recombination (HR) through dephosphorylation of tyrosine 315 on RAD51.Nucleic Acids Res. 2024 Feb 9;52(3):1173-1187. doi: 10.1093/nar/gkad1177. Nucleic Acids Res. 2024. PMID: 38084915 Free PMC article.
-
Single-molecule localization microscopy reveals molecular transactions during RAD51 filament assembly at cellular DNA damage sites.Nucleic Acids Res. 2018 Mar 16;46(5):2398-2416. doi: 10.1093/nar/gkx1303. Nucleic Acids Res. 2018. PMID: 29309696 Free PMC article.
-
RPA and RAD51: fork reversal, fork protection, and genome stability.Nat Struct Mol Biol. 2018 Jun;25(6):446-453. doi: 10.1038/s41594-018-0075-z. Epub 2018 May 28. Nat Struct Mol Biol. 2018. PMID: 29807999 Free PMC article. Review.
-
Molecular flexibility of DNA as a key determinant of RAD51 recruitment.EMBO J. 2020 Apr 1;39(7):e103002. doi: 10.15252/embj.2019103002. Epub 2020 Jan 14. EMBO J. 2020. PMID: 31943278 Free PMC article.
References
-
- Abràmoff M.D., Magalhães P.J., and Ram S.J.. 2004. Image processing with ImageJ. Biophotonics International. 11:36–42.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
