

The oral lipid sensor GPR120 is not indispensable for the orosensory detection of dietary lipids in mice
- PMID: 25489006
- PMCID: PMC4306690
- DOI: 10.1194/jlr.M055202
The oral lipid sensor GPR120 is not indispensable for the orosensory detection of dietary lipids in mice
Abstract
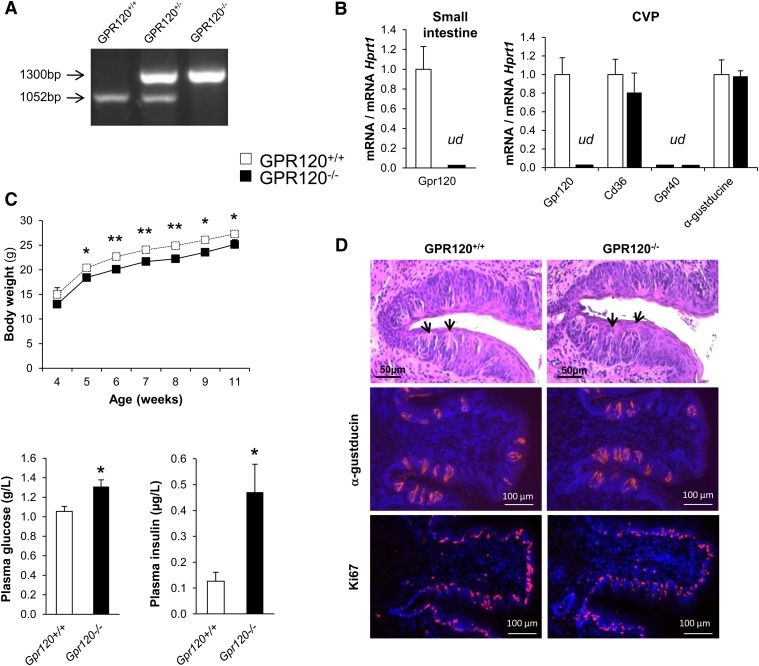
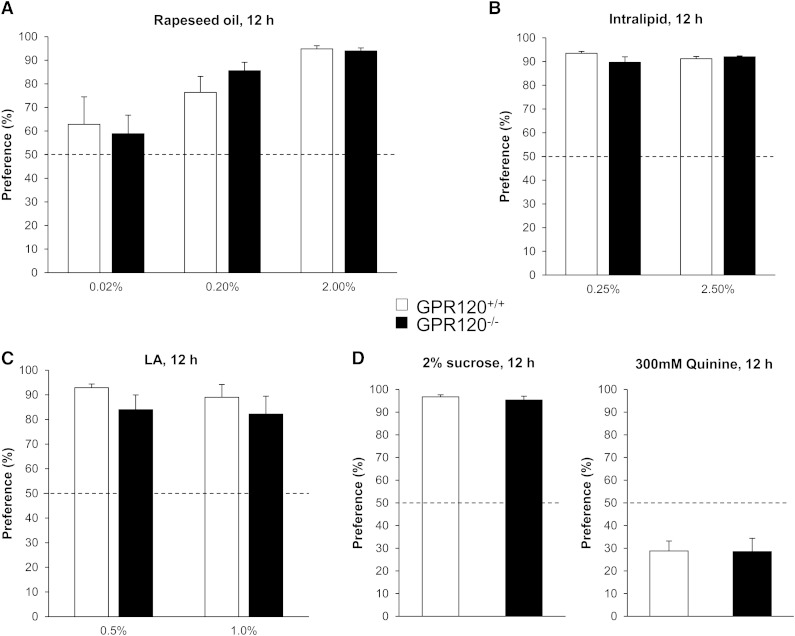
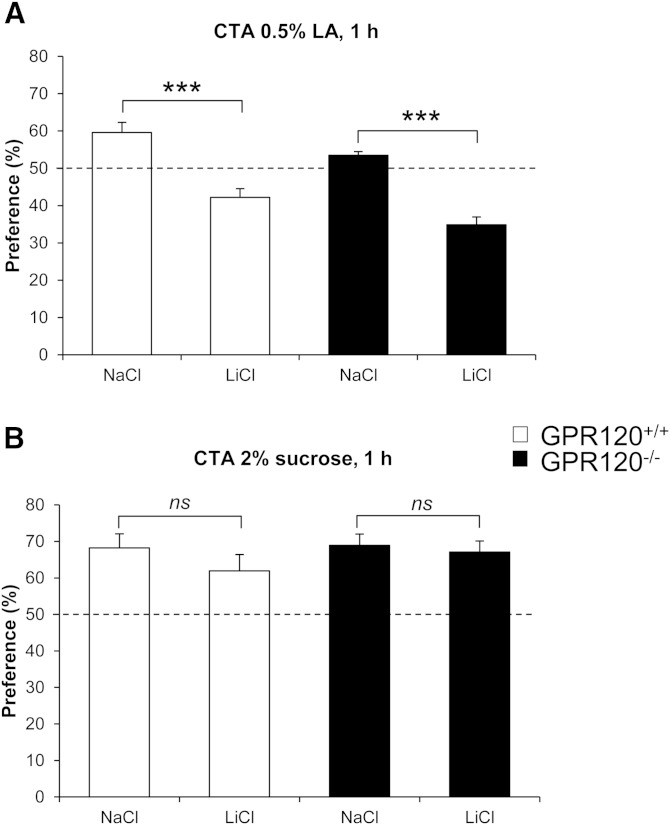
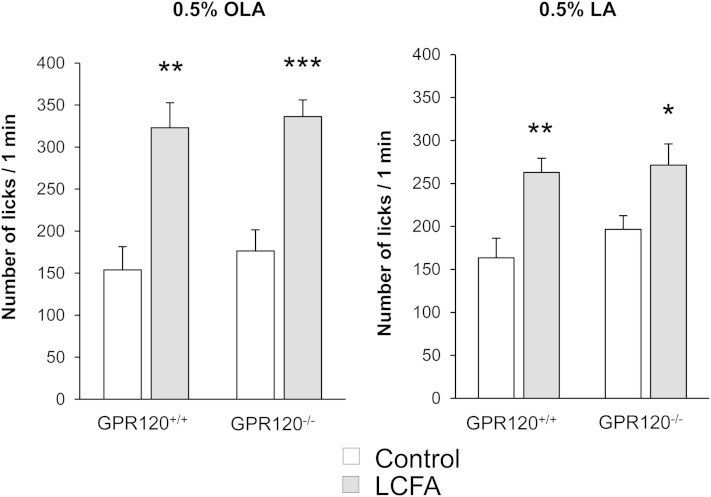
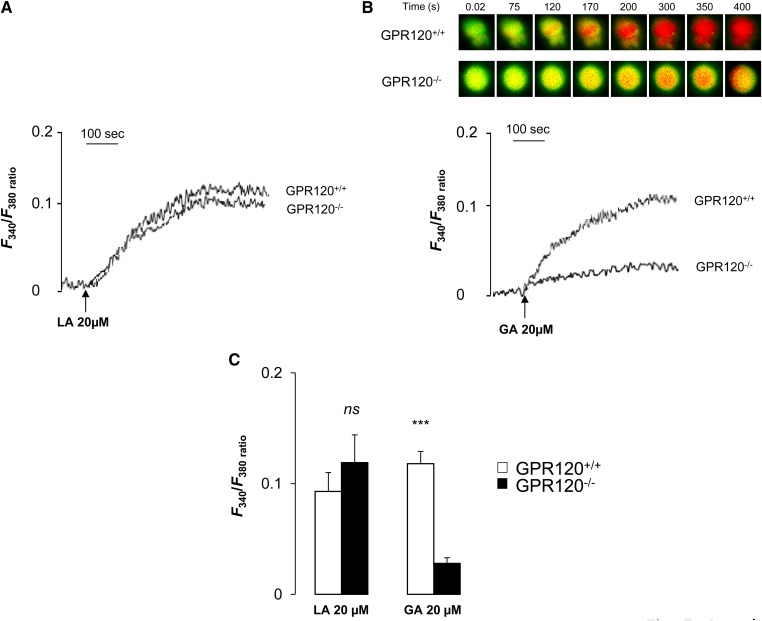
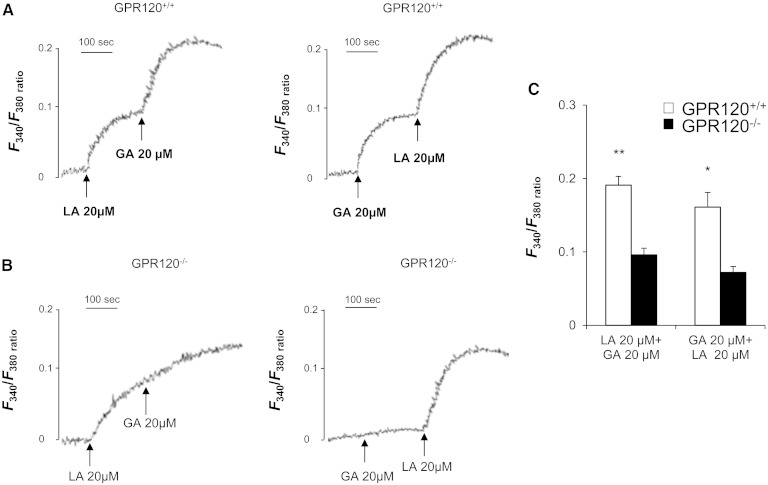
Implication of the long-chain fatty acid (LCFA) receptor GPR120, also termed free fatty acid receptor 4, in the taste-guided preference for lipids is a matter of debate. To further unravel the role of GPR120 in the "taste of fat", the present study was conducted on GPR120-null mice and their wild-type littermates. Using a combination of morphological [i.e., immunohistochemical staining of circumvallate papillae (CVP)], behavioral (i.e., two-bottle preference tests, licking tests and conditioned taste aversion) and functional studies [i.e., calcium imaging in freshly isolated taste bud cells (TBCs)], we show that absence of GPR120 in the oral cavity was not associated with changes in i) gross anatomy of CVP, ii) LCFA-mediated increases in intracellular calcium levels ([Ca(2+)]i), iii) preference for oily and LCFA solutions and iv) conditioned avoidance of LCFA solutions. In contrast, the rise in [Ca(2+)]i triggered by grifolic acid, a specific GPR120 agonist, was dramatically curtailed when the GPR120 gene was lacking. Taken together, these data demonstrate that activation of lingual GPR120 and preference for fat are not connected, suggesting that GPR120 expressed in TBCs is not absolutely required for oral fat detection in mice.
Keywords: G-protein; diet and dietary lipids; fat taste; feeding behavior; lipids; mouse; nutrition; receptors.
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Kawai T., Fushiki T. 2003. Importance of lipolysis in oral cavity for orosensory detection of fat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 285: R447–R454. - PubMed
-
- Baillie A. G., Coburn C. T., Abumrad N. A. 1996. Reversible binding of long-chain fatty acids to purified FAT, the adipose CD36 homolog. J. Membr. Biol. 153: 75–81. - PubMed
-
- Fukuwatari T., Kawada T., Tsuruta M., Hiraoka T., Iwanaga T., Sugimoto E., Fushiki T. 1997. Expression of the putative membrane fatty acid transporter (FAT) in taste buds of the circumvallate papillae in rats. FEBS Lett. 414: 461–464. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
