

A novel manganese-dependent ATM-p53 signaling pathway is selectively impaired in patient-based neuroprogenitor and murine striatal models of Huntington's disease
- PMID: 25489053
- PMCID: PMC4355025
- DOI: 10.1093/hmg/ddu609
A novel manganese-dependent ATM-p53 signaling pathway is selectively impaired in patient-based neuroprogenitor and murine striatal models of Huntington's disease
Abstract
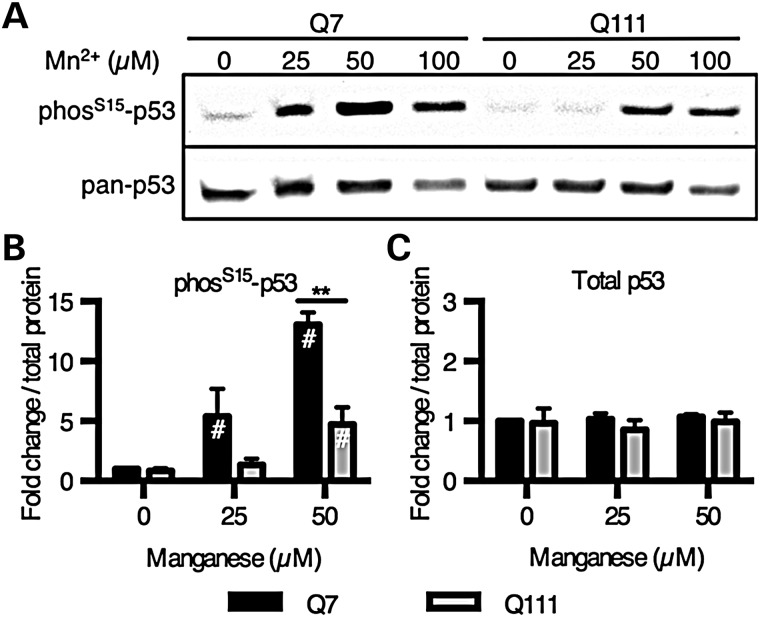
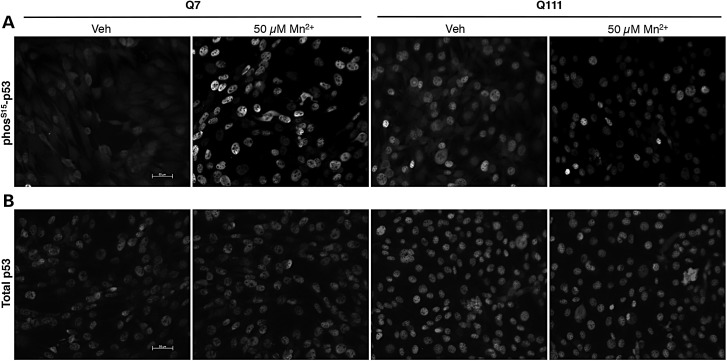
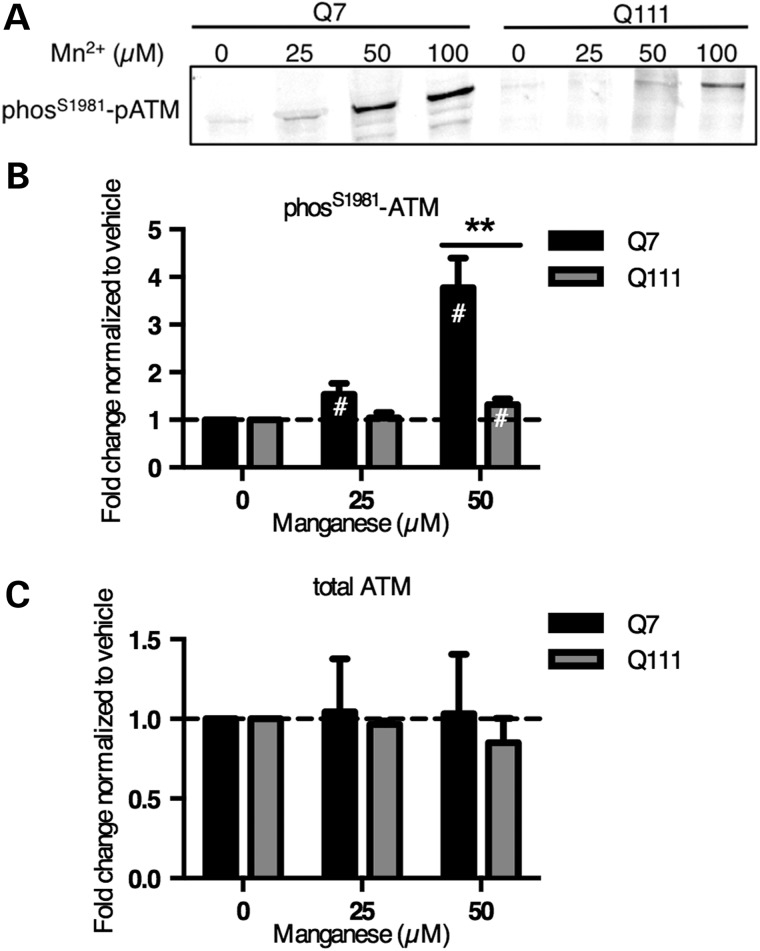
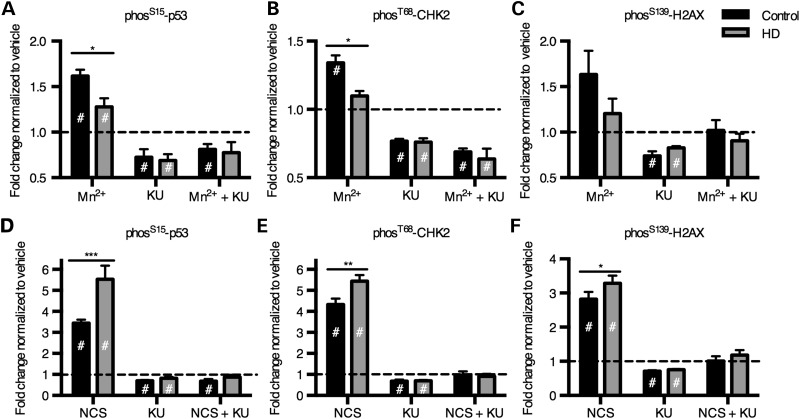
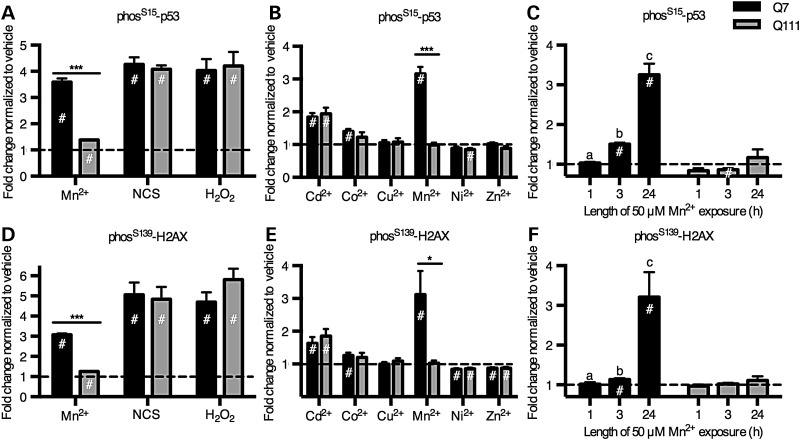
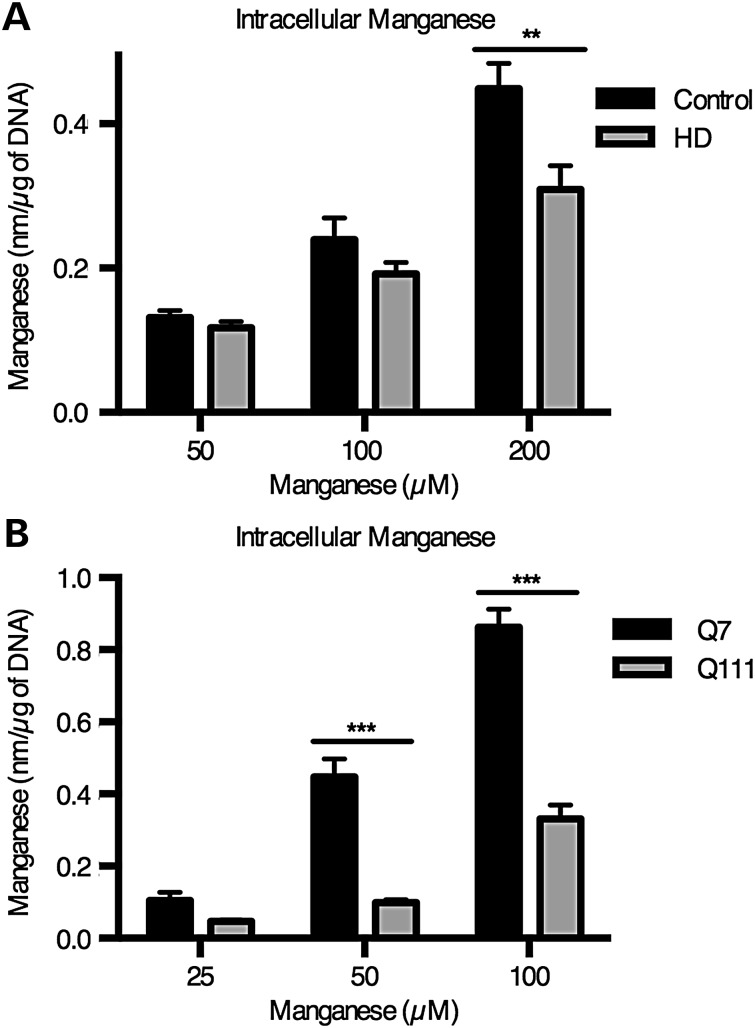
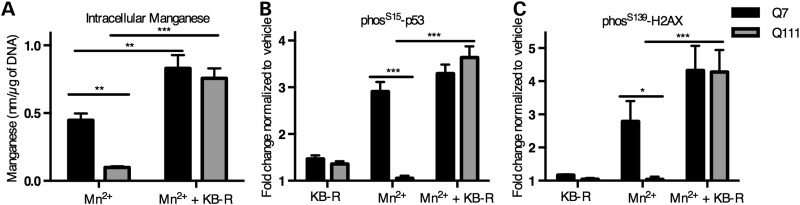
The essential micronutrient manganese is enriched in brain, especially in the basal ganglia. We sought to identify neuronal signaling pathways responsive to neurologically relevant manganese levels, as previous data suggested that alterations in striatal manganese handling occur in Huntington's disease (HD) models. We found that p53 phosphorylation at serine 15 is the most responsive cell signaling event to manganese exposure (of 18 tested) in human neuroprogenitors and a mouse striatal cell line. Manganese-dependent activation of p53 was severely diminished in HD cells. Inhibitors of ataxia telangiectasia mutated (ATM) kinase decreased manganese-dependent phosphorylation of p53. Likewise, analysis of ATM autophosphorylation and additional ATM kinase targets, H2AX and CHK2, support a role for ATM in the activation of p53 by manganese and that a defect in this process occurs in HD. Furthermore, the deficit in Mn-dependent activation of ATM kinase in HD neuroprogenitors was highly selective, as DNA damage and oxidative injury, canonical activators of ATM, did not show similar deficits. We assessed cellular manganese handling to test for correlations with the ATM-p53 pathway, and we observed reduced Mn accumulation in HD human neuroprogenitors and HD mouse striatal cells at manganese exposures associated with altered p53 activation. To determine if this phenotype contributes to the deficit in manganese-dependent ATM activation, we used pharmacological manipulation to equalize manganese levels between HD and control mouse striatal cells and rescued the ATM-p53 signaling deficit. Collectively, our data demonstrate selective alterations in manganese biology in cellular models of HD manifest in ATM-p53 signaling.
© The Author 2014. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
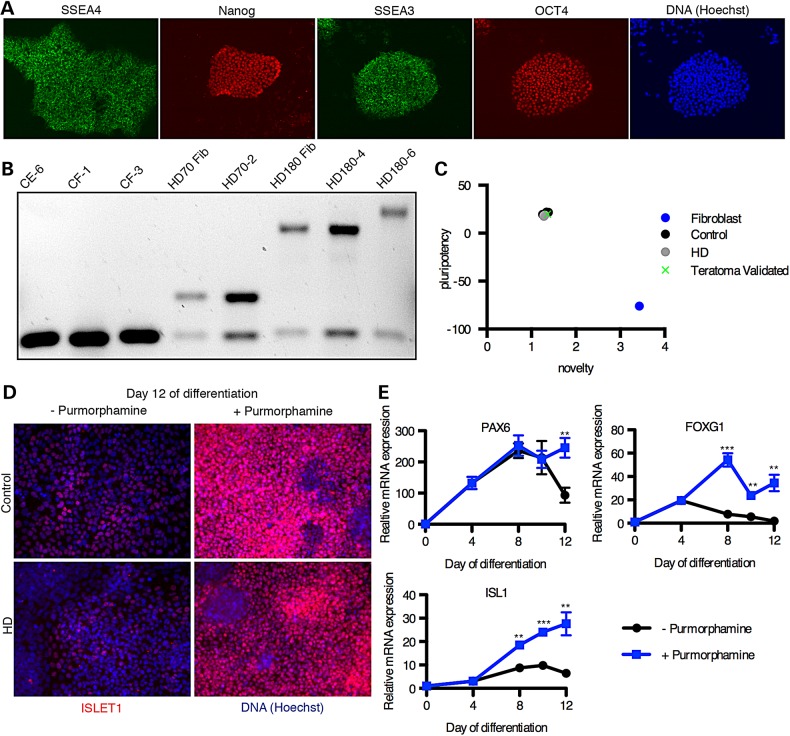
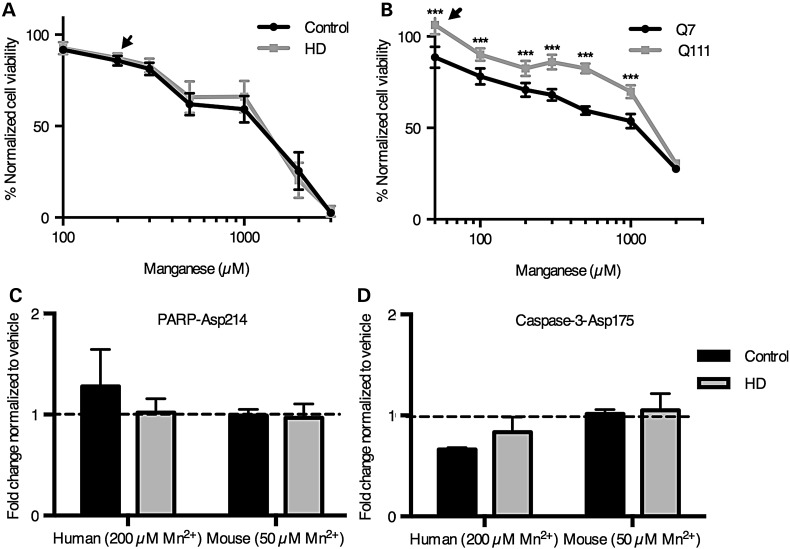
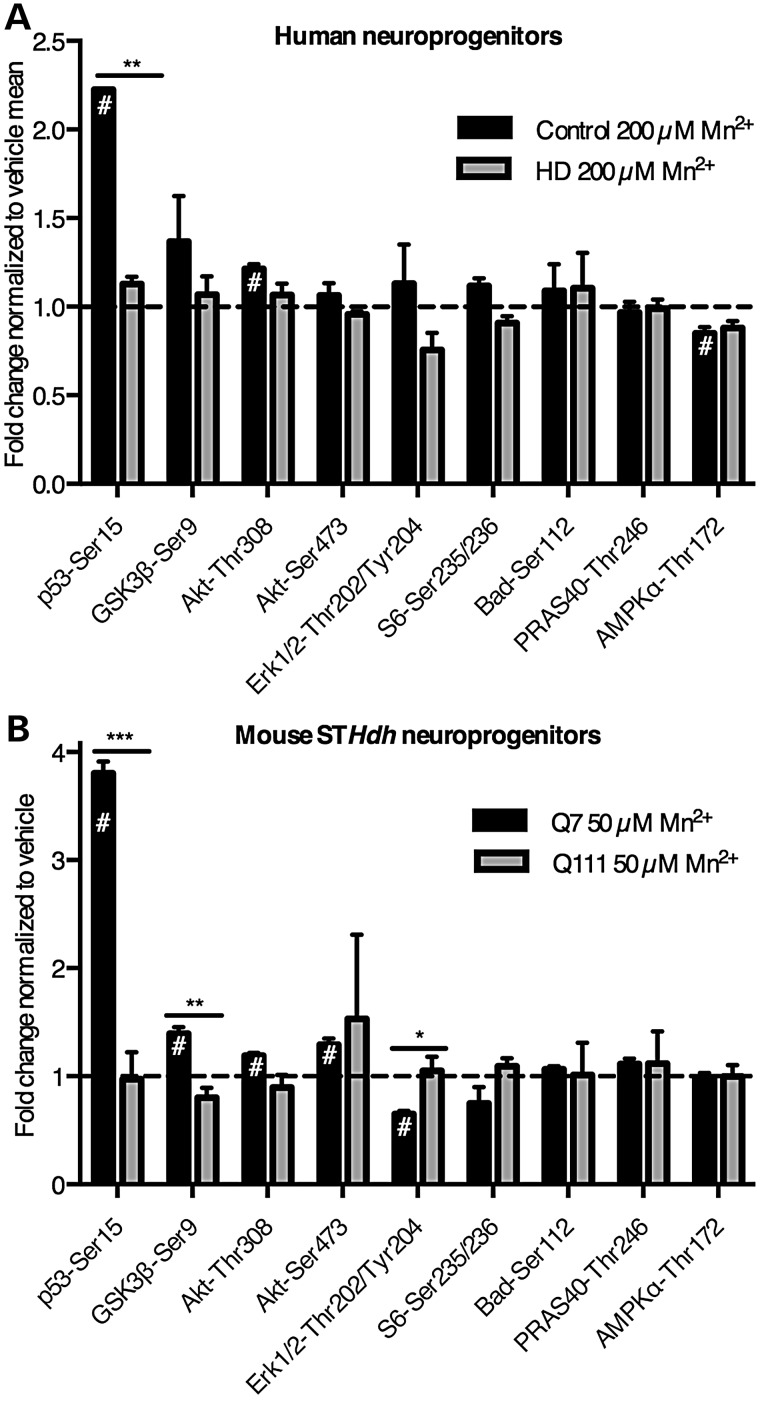
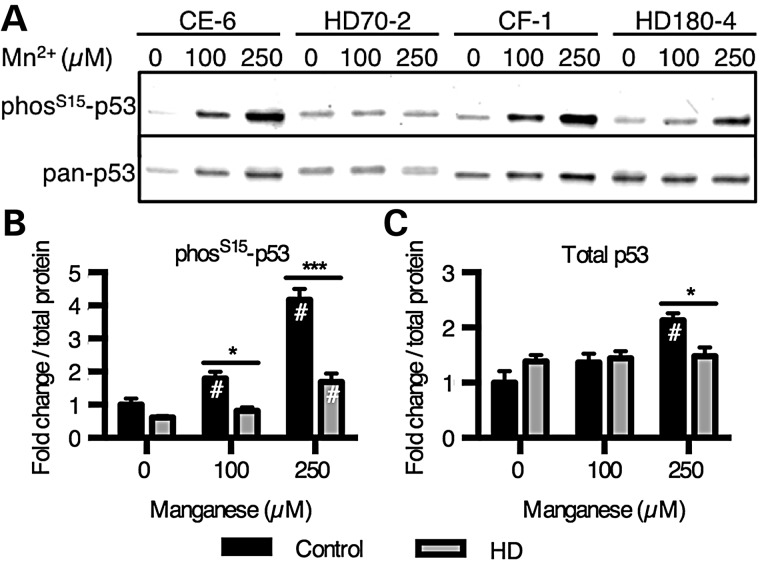
References
-
- Zuccato C., Valenza M., Cattaneo E. Molecular mechanisms and potential therapeutical targets in Huntington’s disease. Physiol. Rev. 2010;90:905–981. - PubMed
-
- Georgiou N., Bradshaw J.L., Chiu E., Tudor A., O'Gorman L., Phillips J.G. Differential clinical and motor control function in a pair of monozygotic twins with Huntington’s disease. Mov. Disord. 1999;14:320–325. - PubMed
-
- Anca M., Gazit E., Loewenthal R., Ostrovsky O., Frydman M., Giladi N. Different phenotypic expression in monozygotic twins with Huntington disease. Am. J. Med. Genet. A. 2004;124:89–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
