

Microtubules regulate focal adhesion dynamics through MAP4K4
- PMID: 25490267
- PMCID: PMC4261153
- DOI: 10.1016/j.devcel.2014.10.025
Microtubules regulate focal adhesion dynamics through MAP4K4
Abstract
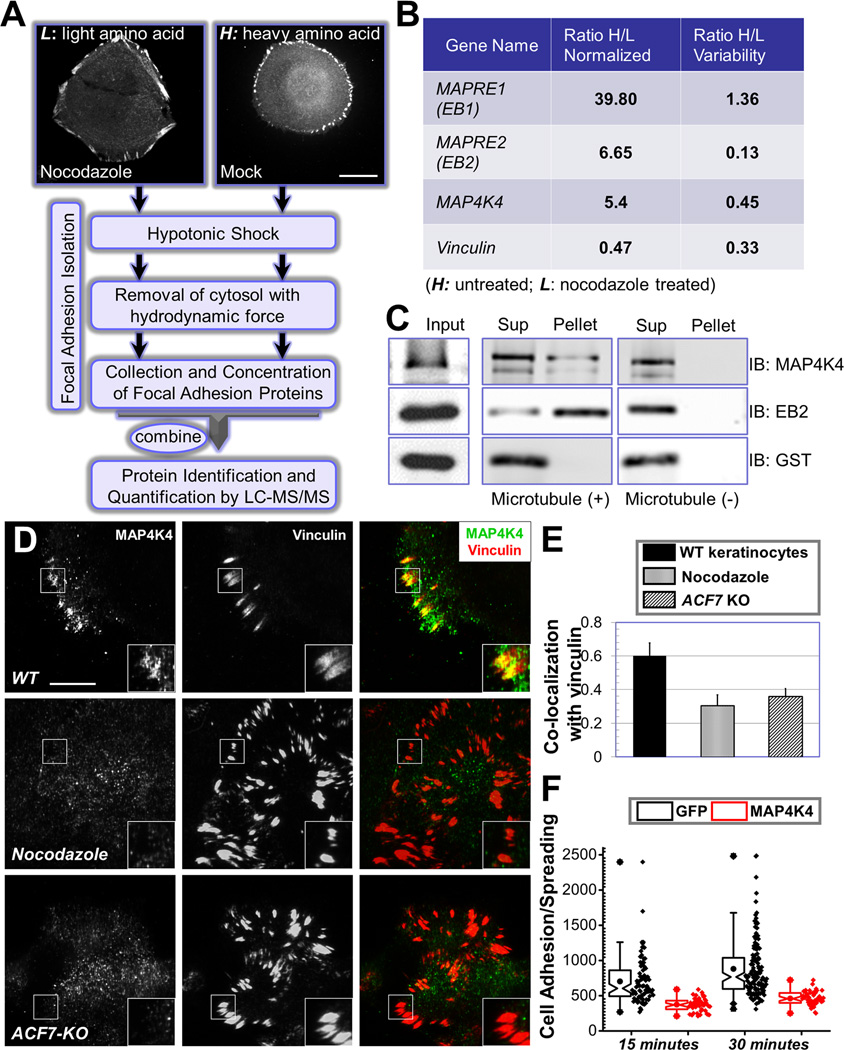
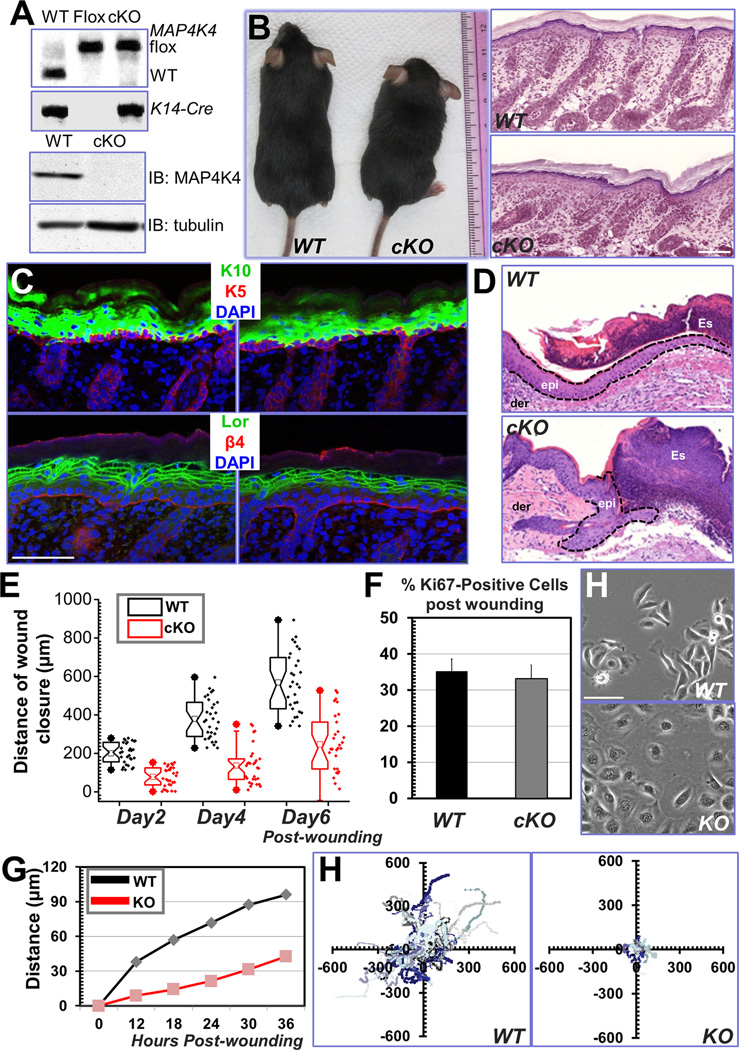
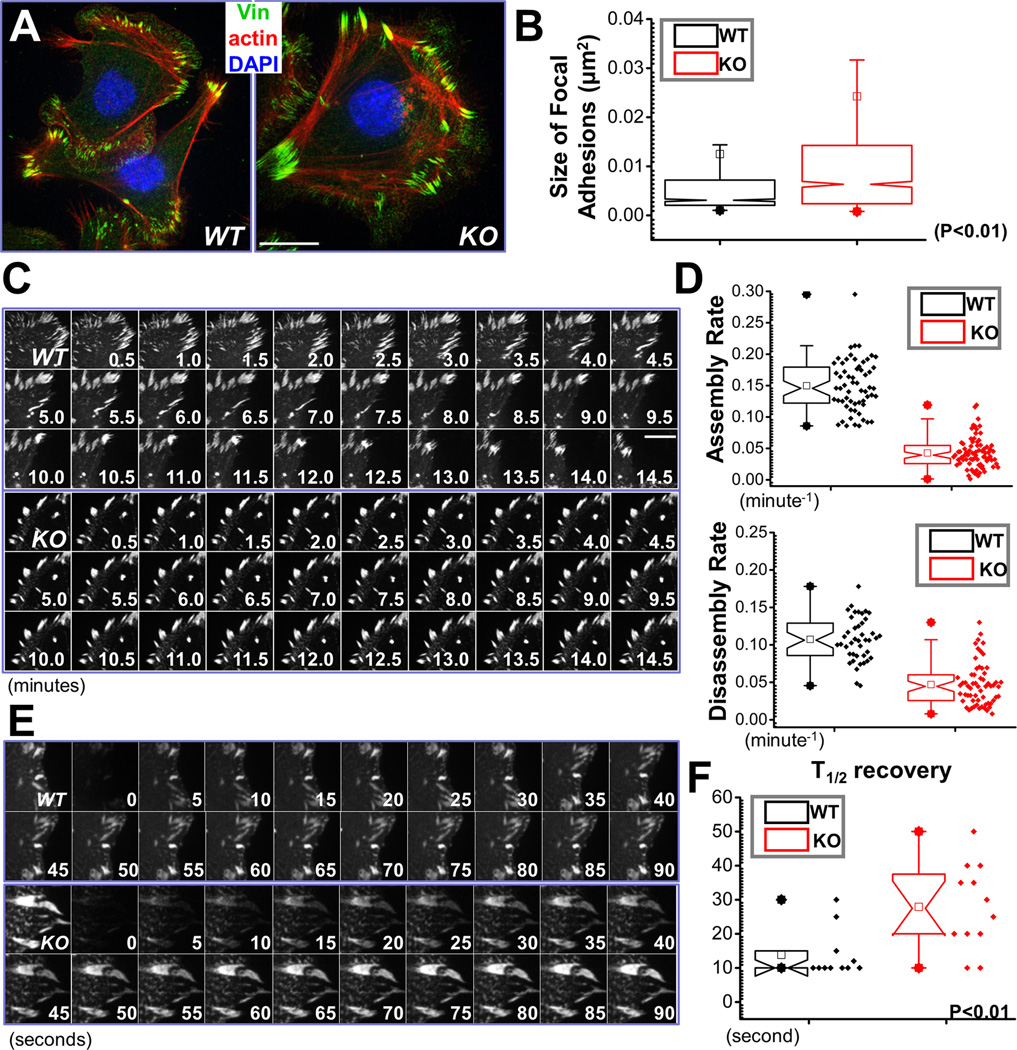
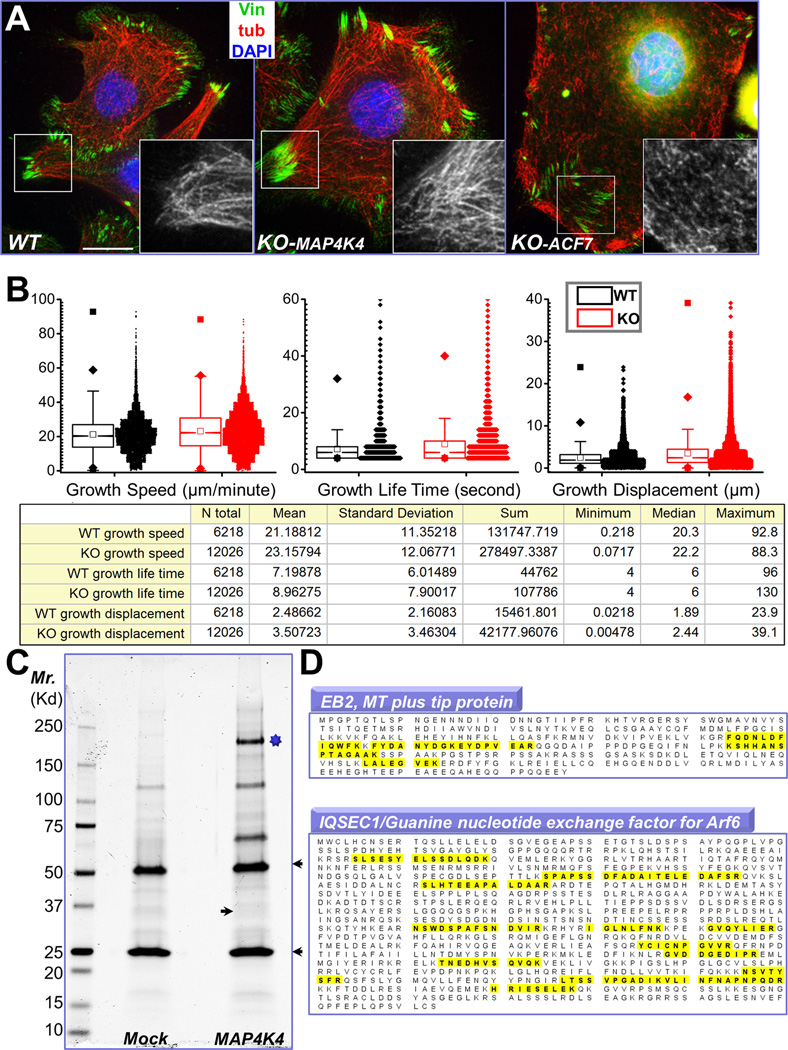
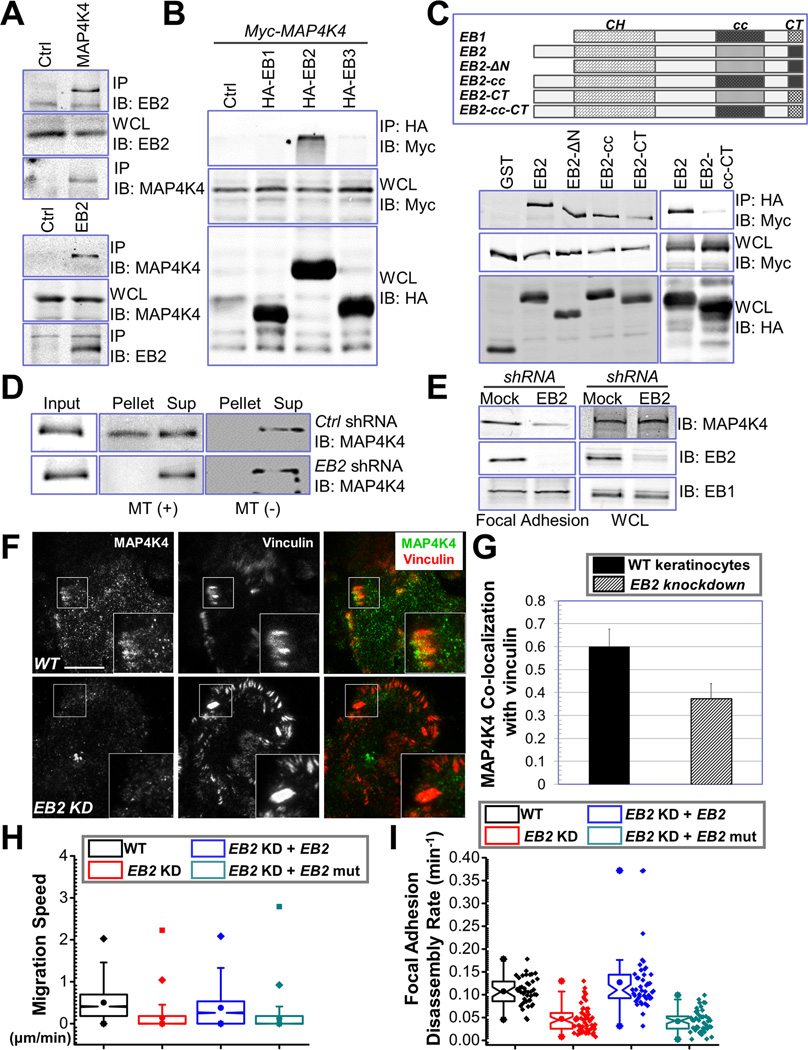
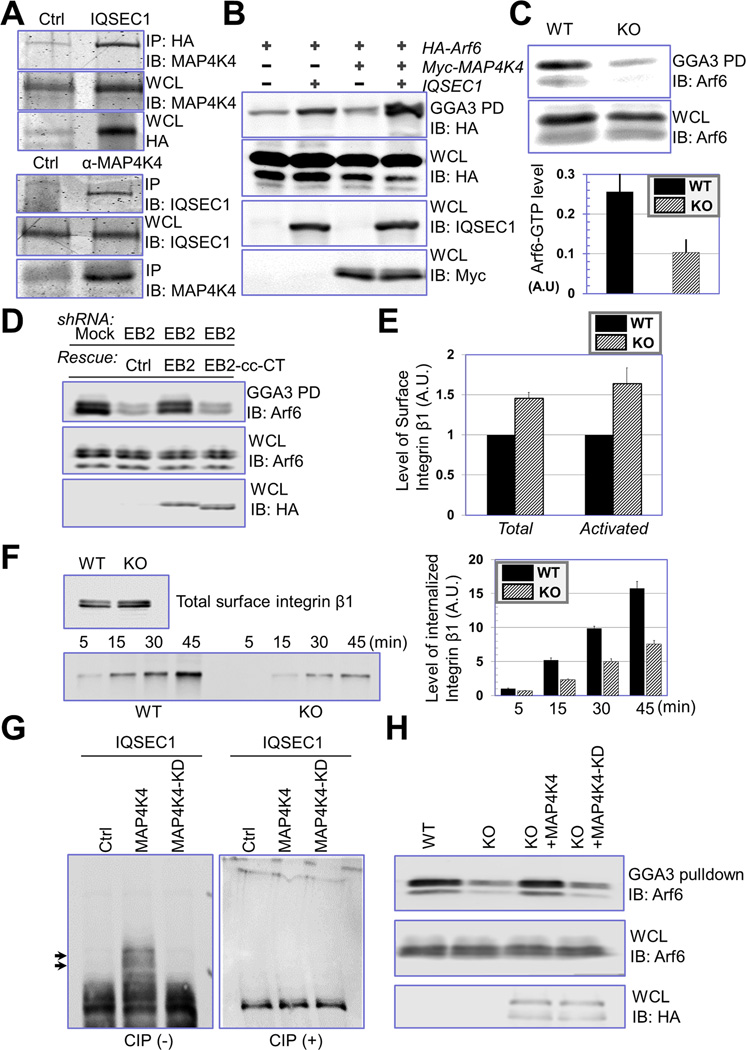
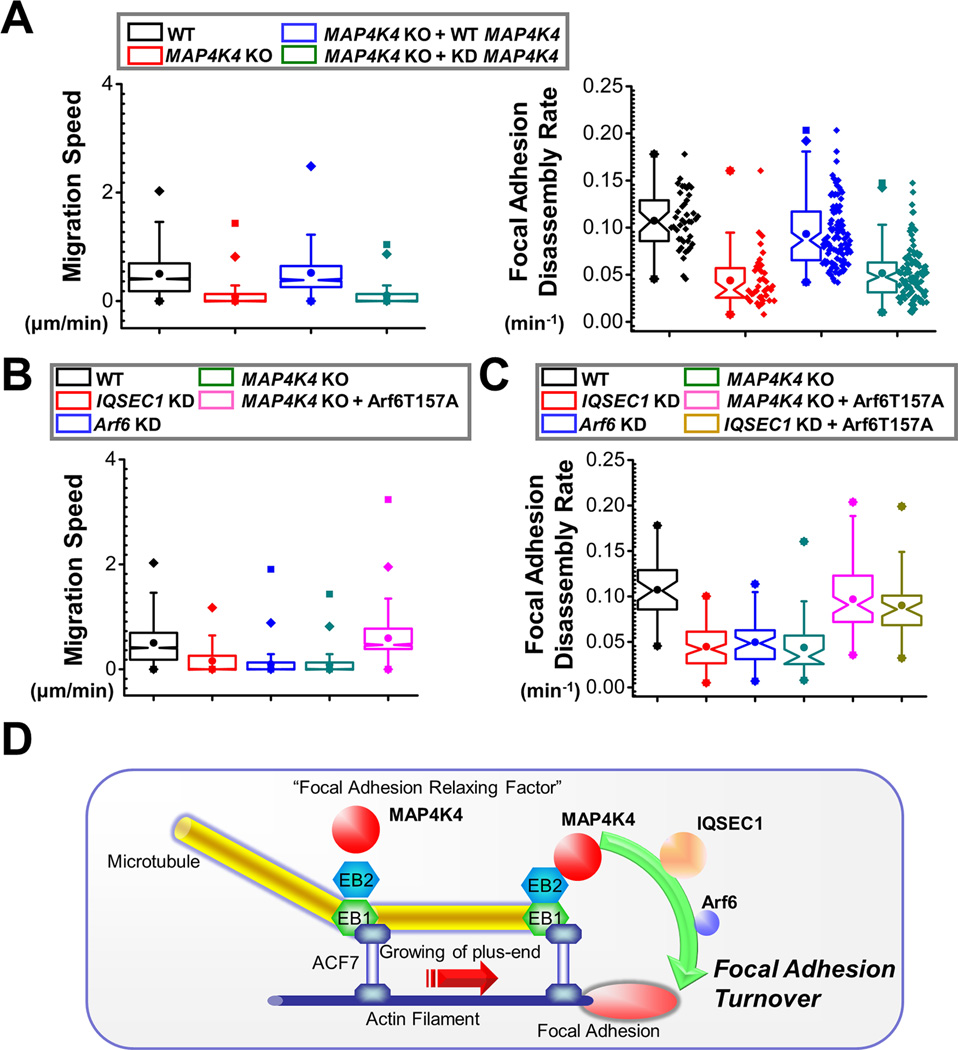
Disassembly of focal adhesions (FAs) allows cell retraction and integrin detachment from the extracellular matrix, processes critical for cell movement. Growth of microtubules (MTs) can promote FA turnover by serving as tracks to deliver proteins essential for FA disassembly. The molecular nature of this FA "disassembly factor," however, remains elusive. By quantitative proteomics, we identified mitogen-activated protein kinase kinase kinase kinase 4 (MAP4K4) as an FA regulator that associates with MTs. Knockout of MAP4K4 stabilizes FAs and impairs cell migration. By exploring underlying mechanisms, we further show that MAP4K4 associates with ending binding 2 (EB2) and IQ motif and SEC7 domain-containing protein 1 (IQSEC1), a guanine nucleotide exchange factor specific for Arf6, whose activation promotes integrin internalization. Together, our findings provide critical insight into FA disassembly, suggesting that MTs can deliver MAP4K4 toward FAs through EB2, where MAP4K4 can, in turn, activate Arf6 via IQSEC1 and enhance FA dissolution.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Akhmanova A, Steinmetz MO. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nature reviews Molecular cell biology. 2008;9:309–322. - PubMed
-
- Bershadsky A, Chausovsky A, Becker E, Lyubimova A, Geiger B. Involvement of microtubules in the control of adhesion-dependent signal transduction. Current biology : CB. 1996;6:1279–1289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
