

The Arabidopsis cellulose synthase complex: a proposed hexamer of CESA trimers in an equimolar stoichiometry
- PMID: 25490917
- PMCID: PMC4311198
- DOI: 10.1105/tpc.114.131193
The Arabidopsis cellulose synthase complex: a proposed hexamer of CESA trimers in an equimolar stoichiometry
Abstract
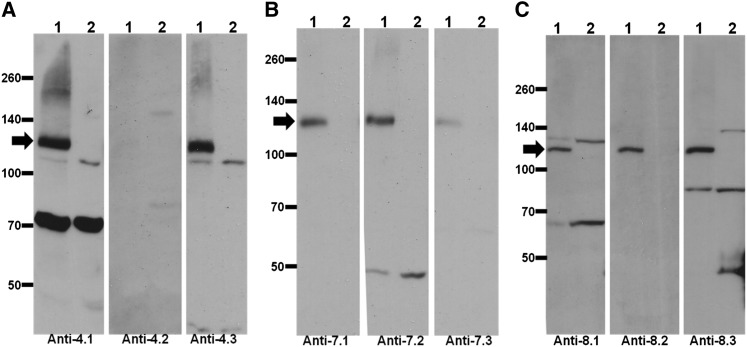
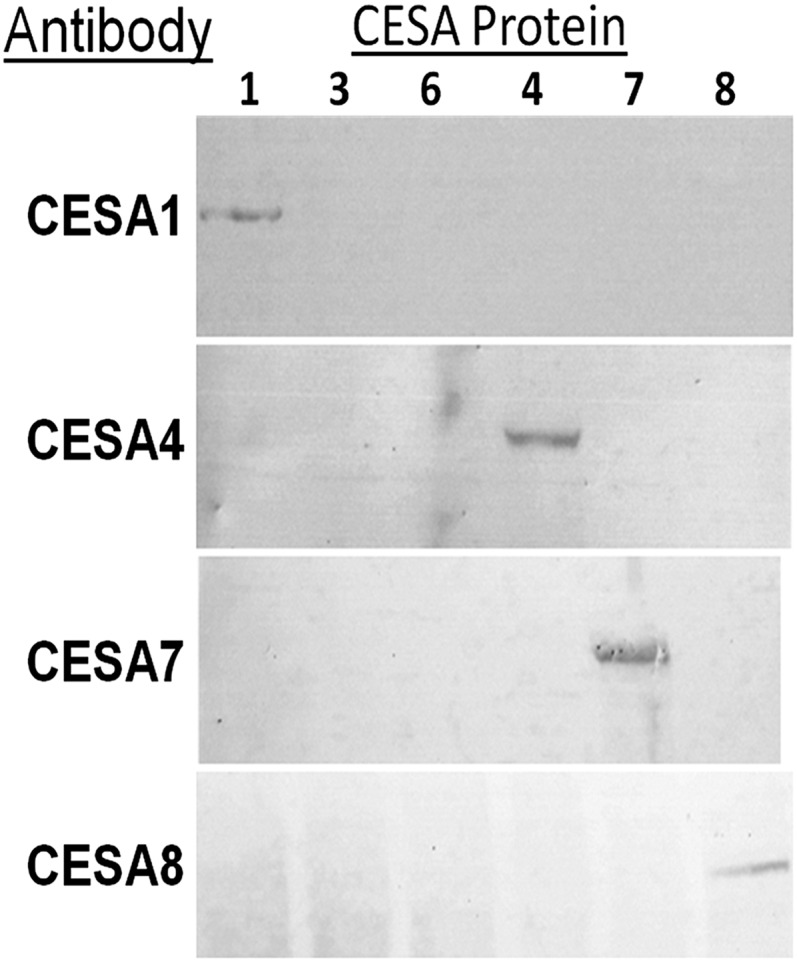
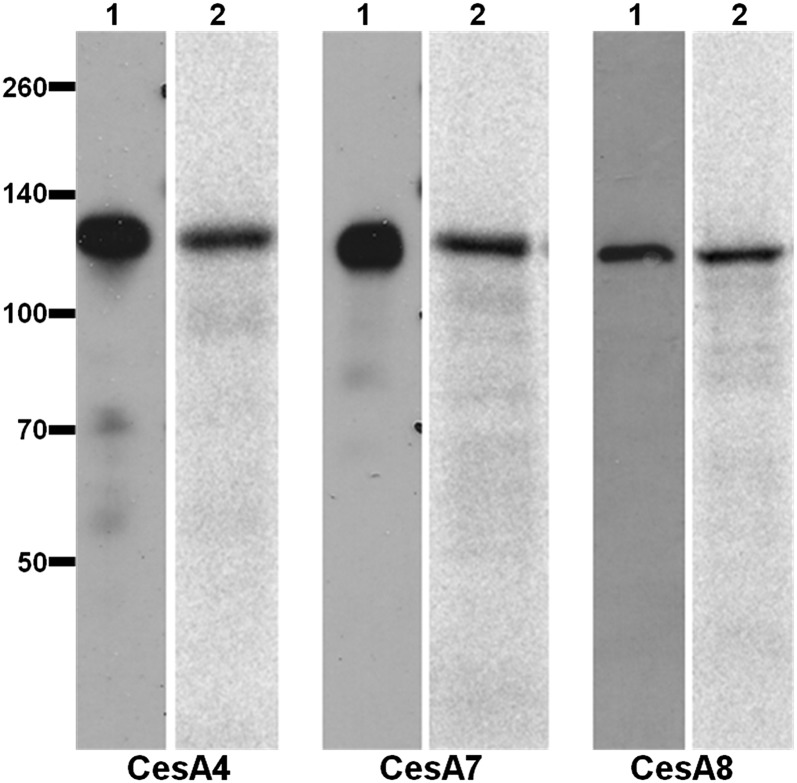
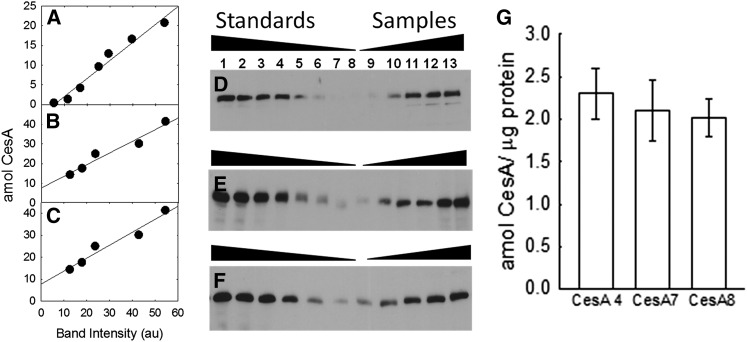
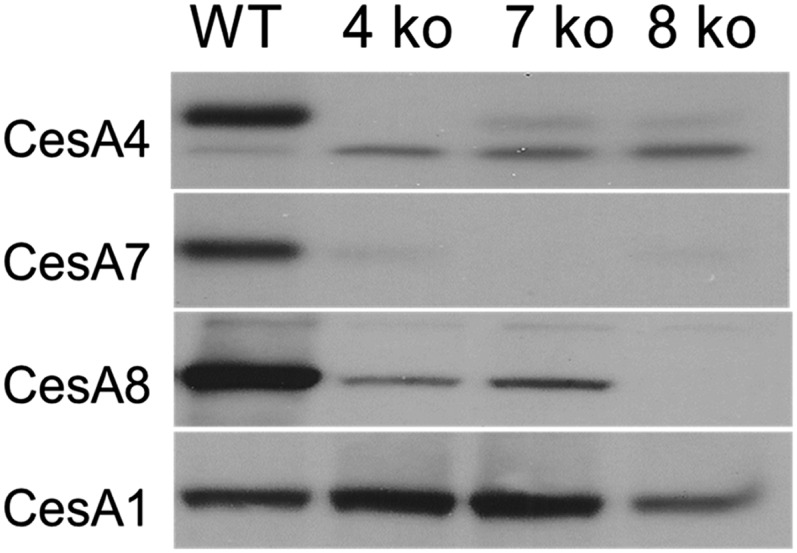
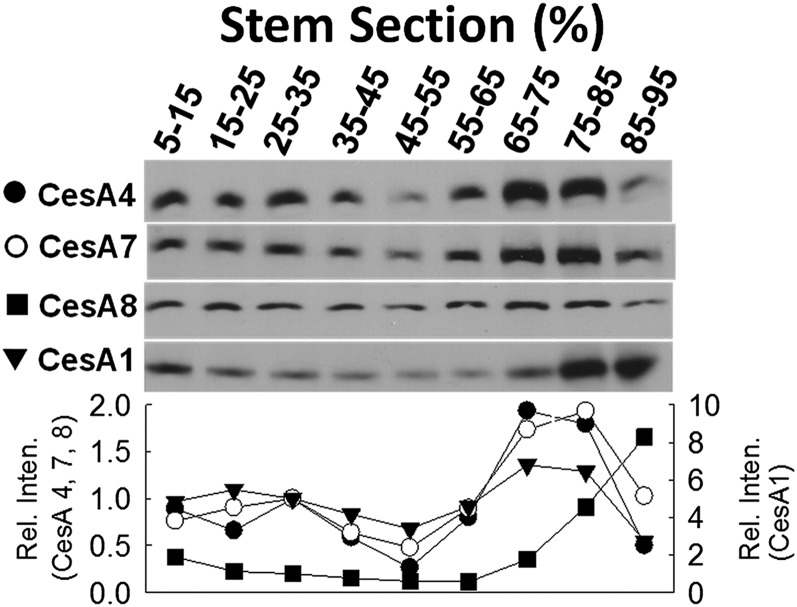
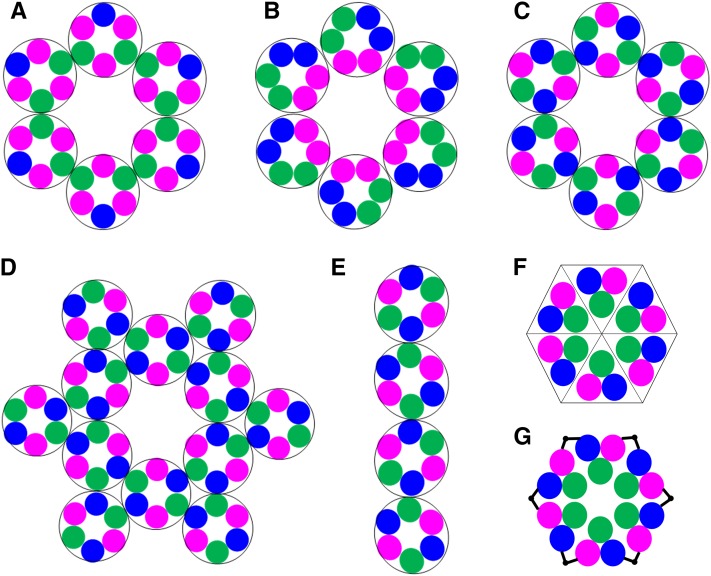
Cellulose is the most abundant renewable polymer on Earth and a major component of the plant cell wall. In vascular plants, cellulose synthesis is catalyzed by a large, plasma membrane-localized cellulose synthase complex (CSC), visualized as a hexameric rosette structure. Three unique cellulose synthase (CESA) isoforms are required for CSC assembly and function. However, elucidation of either the number or stoichiometry of CESAs within the CSC has remained elusive. In this study, we show a 1:1:1 stoichiometry between the three Arabidopsis thaliana secondary cell wall isozymes: CESA4, CESA7, and CESA8. This ratio was determined utilizing a simple but elegant method of quantitative immunoblotting using isoform-specific antibodies and (35)S-labeled protein standards for each CESA. Additionally, the observed equimolar stoichiometry was found to be fixed along the axis of the stem, which represents a developmental gradient. Our results complement recent spectroscopic analyses pointing toward an 18-chain cellulose microfibril. Taken together, we propose that the CSC is composed of a hexamer of catalytically active CESA trimers, with each CESA in equimolar amounts. This finding is a crucial advance in understanding how CESAs integrate to form higher order complexes, which is a key determinate of cellulose microfibril and cell wall properties.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
-
- Atanassov I.I., Pittman J.K., Turner S.R. (2009). Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J. Biol. Chem. 284: 3833–3841. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
