

Small RNA profiling reveals regulation of Arabidopsis miR168 and heterochromatic siRNA415 in response to fungal elicitors
- PMID: 25491154
- PMCID: PMC4299684
- DOI: 10.1186/1471-2164-15-1083
Small RNA profiling reveals regulation of Arabidopsis miR168 and heterochromatic siRNA415 in response to fungal elicitors
Abstract
Background: Small RNAs (sRNAs), including small interfering RNAs (siRNAs) and microRNAs (miRNAs), have emerged as important regulators of eukaryotic gene expression. In plants, miRNAs play critical roles in development, nutrient homeostasis and abiotic stress responses. Accumulating evidence also reveals that sRNAs are involved in plant immunity. Most studies on pathogen-regulated sRNAs have been conducted in Arabidopsis plants infected with the bacterial pathogen Pseudomonas syringae, or treated with the flagelin-derived elicitor peptide flg22 from P. syringae. This work investigates sRNAs that are regulated by elicitors from the fungus Fusarium oxysporum in Arabidopsis.
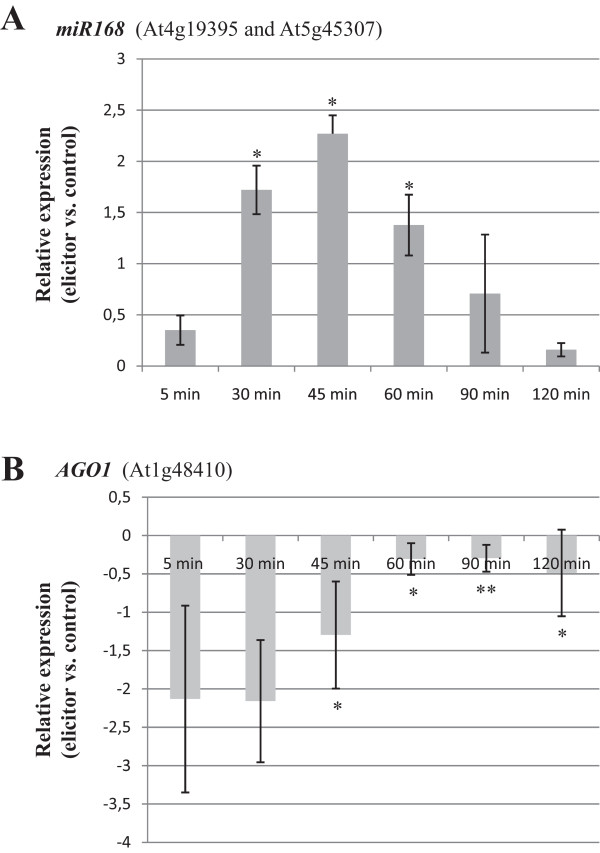
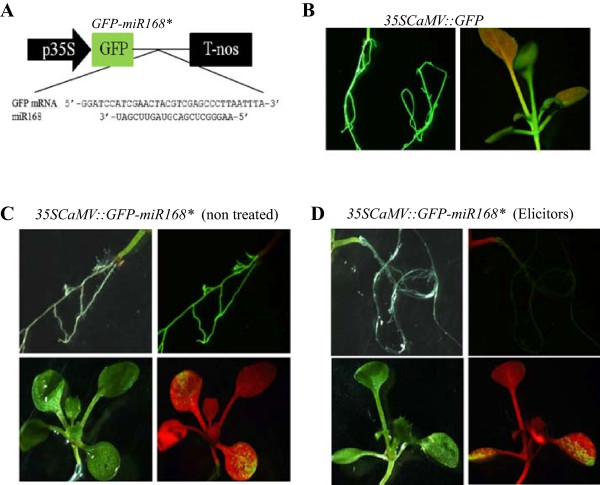
Results: Microarray analysis revealed alterations on the accumulation of a set of sRNAs in response to elicitor treatment, including miRNAs and small RNA sequences derived from massively parallel signature sequencing. Among the elicitor-regulated miRNAs was miR168 which regulates ARGONAUTE1, the core component of the RNA-induced silencing complex involved in miRNA functioning. Promoter analysis in transgenic Arabidopsis plants revealed transcriptional activation of MIR168 by fungal elicitors. Furthermore, transgenic plants expressing a GFP-miR168 sensor gene confirmed that the elicitor-induced miR168 is active. MiR823, targeting Chromomethylase3 (CMT3) involved in RNA-directed DNA methylation (RdDM) was also found to be regulated by fungal elicitors. In addition to known miRNAs, microarray analysis allowed the identification of an elicitor-inducible small RNA that was incorrectly annotated as a miRNA. Studies on Arabidopsis mutants impaired in small RNA biogenesis demonstrated that this sRNA, is a heterochromatic-siRNA (hc-siRNA) named as siRNA415. Hc-siRNAs are known to be involved in RNA-directed DNA methylation (RdDM). SiRNA415 is detected in several plant species.
Conclusion: Results here presented support a transcriptional regulatory mechanism underlying MIR168 expression. This finding highlights the importance of miRNA functioning in adaptive processes of Arabidopsis plants to fungal infection. The results of this study also lay a foundation for the involvement of RdDM processes through the activity of siRNA415 and miR823 in mediating regulation of immune responses in Arabidopsis plants.
Figures



Similar articles
-
MicroRNA-mediated regulation of gene expression in the response of rice plants to fungal elicitors.RNA Biol. 2015;12(8):847-63. doi: 10.1080/15476286.2015.1050577. RNA Biol. 2015. PMID: 26083154 Free PMC article.
-
Identification of Arabidopsis thaliana small RNAs responsive to the fungal pathogen Botrytis cinerea at an early stage of interaction.PLoS One. 2024 Jun 14;19(6):e0304790. doi: 10.1371/journal.pone.0304790. eCollection 2024. PLoS One. 2024. PMID: 38875250 Free PMC article.
-
Specific impact of tobamovirus infection on the Arabidopsis small RNA profile.PLoS One. 2011 May 10;6(5):e19549. doi: 10.1371/journal.pone.0019549. PLoS One. 2011. PMID: 21572953 Free PMC article.
-
Recent advances in small RNA mediated plant-virus interactions.Crit Rev Biotechnol. 2019 Jun;39(4):587-601. doi: 10.1080/07388551.2019.1597830. Epub 2019 Apr 4. Crit Rev Biotechnol. 2019. PMID: 30947560 Review.
-
Small regulatory RNAs in rice epigenetic regulation.Biochem Soc Trans. 2022 Jun 30;50(3):1215-1225. doi: 10.1042/BST20210336. Biochem Soc Trans. 2022. PMID: 35579290 Review.
Cited by
-
Diverse Functions of Small RNAs in Different Plant-Pathogen Communications.Front Microbiol. 2016 Oct 4;7:1552. doi: 10.3389/fmicb.2016.01552. eCollection 2016. Front Microbiol. 2016. PMID: 27757103 Free PMC article. Review.
-
MicroRNA-mediated regulation of gene expression in the response of rice plants to fungal elicitors.RNA Biol. 2015;12(8):847-63. doi: 10.1080/15476286.2015.1050577. RNA Biol. 2015. PMID: 26083154 Free PMC article.
-
MicroRNA candidate miRcand137 in apple is induced by Botryosphaeria dothidea for impairing host defense.Plant Physiol. 2022 Jun 27;189(3):1814-1832. doi: 10.1093/plphys/kiac171. Plant Physiol. 2022. PMID: 35512059 Free PMC article.
-
Transcriptomic Crosstalk between Fungal Invasive Pathogens and Their Host Cells: Opportunities and Challenges for Next-Generation Sequencing Methods.J Fungi (Basel). 2016 Jan 14;2(1):7. doi: 10.3390/jof2010007. J Fungi (Basel). 2016. PMID: 29376924 Free PMC article. Review.
-
Research Progress on miRNAs and Artificial miRNAs in Insect and Disease Resistance and Breeding in Plants.Genes (Basel). 2024 Sep 12;15(9):1200. doi: 10.3390/genes15091200. Genes (Basel). 2024. PMID: 39336791 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
