

GCN2 is required to increase fibroblast growth factor 21 and maintain hepatic triglyceride homeostasis during asparaginase treatment
- PMID: 25491724
- PMCID: PMC4329494
- DOI: 10.1152/ajpendo.00361.2014
GCN2 is required to increase fibroblast growth factor 21 and maintain hepatic triglyceride homeostasis during asparaginase treatment
Abstract
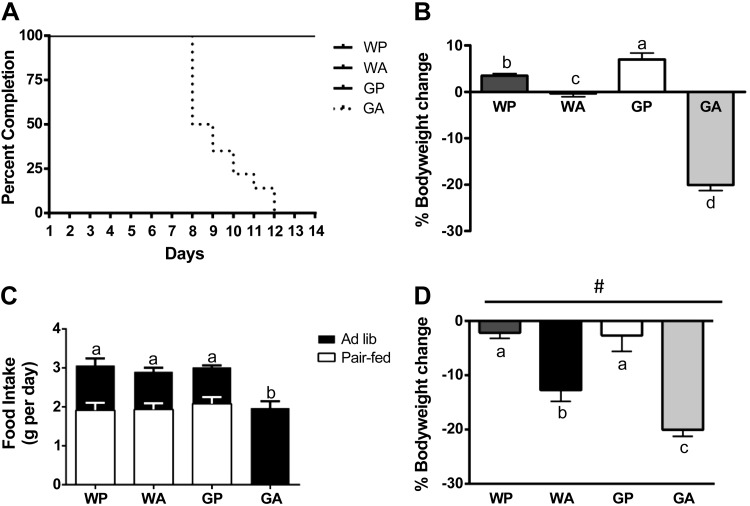
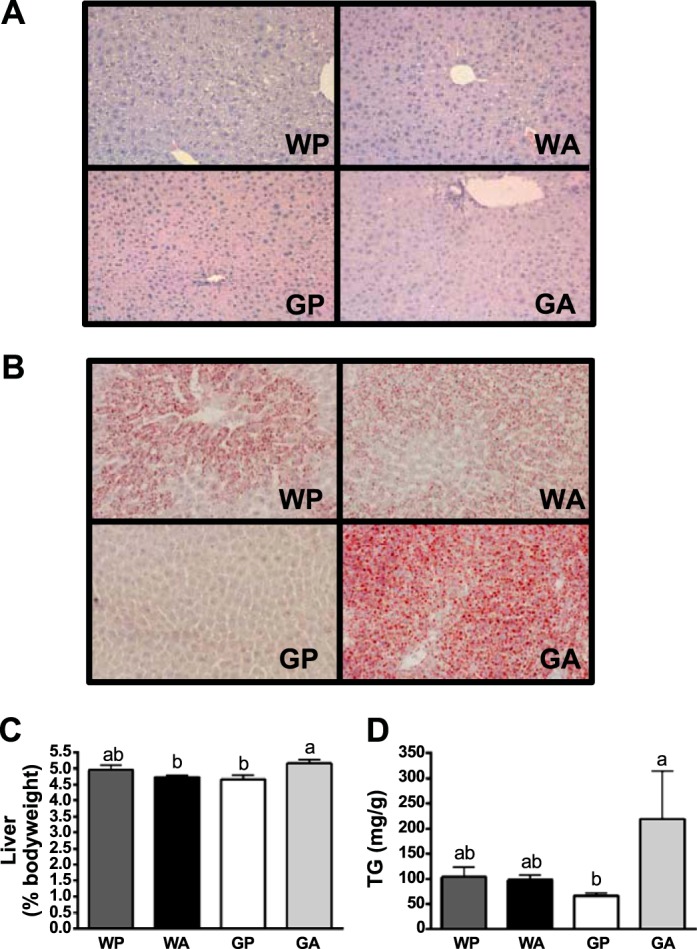
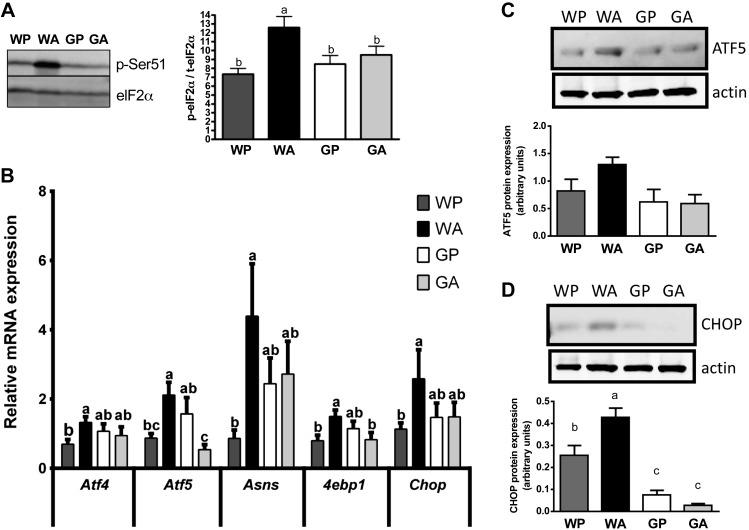
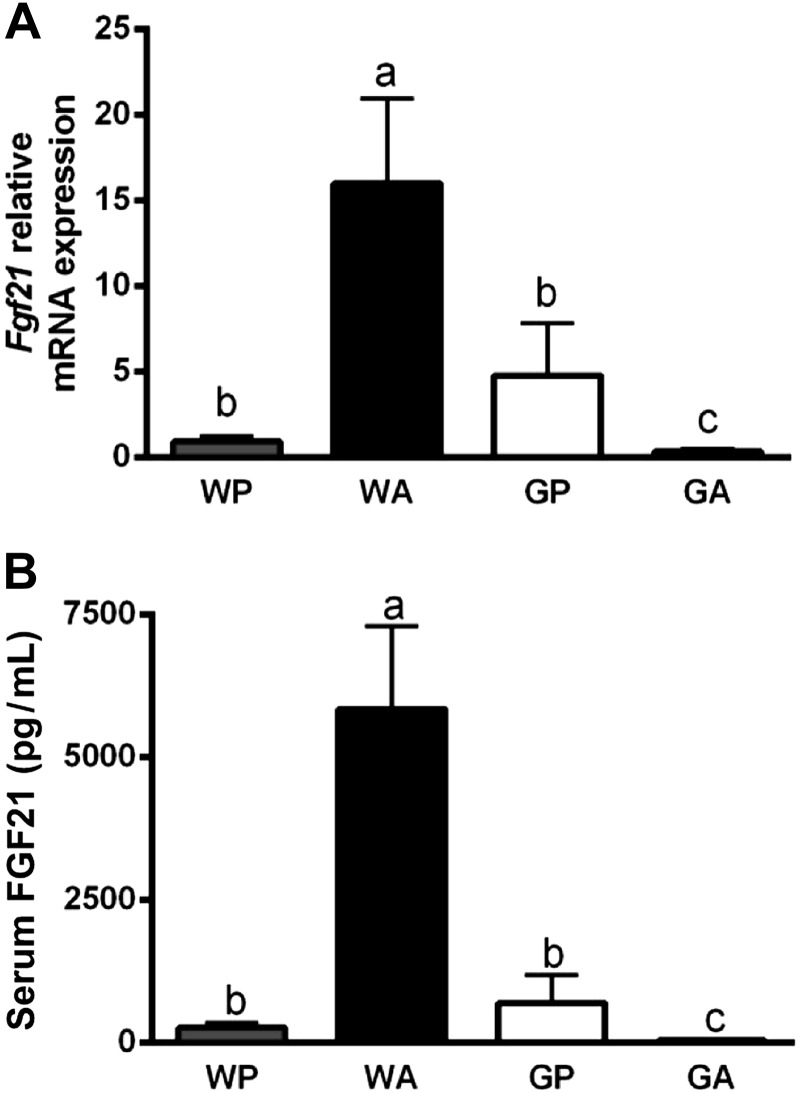
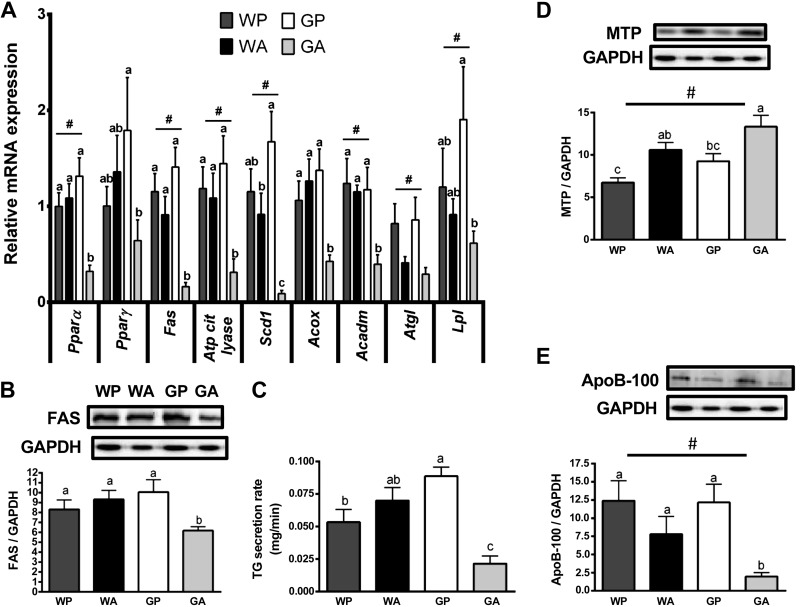
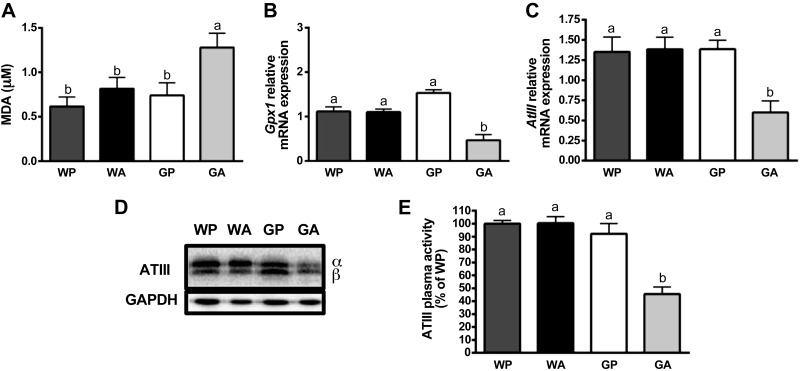
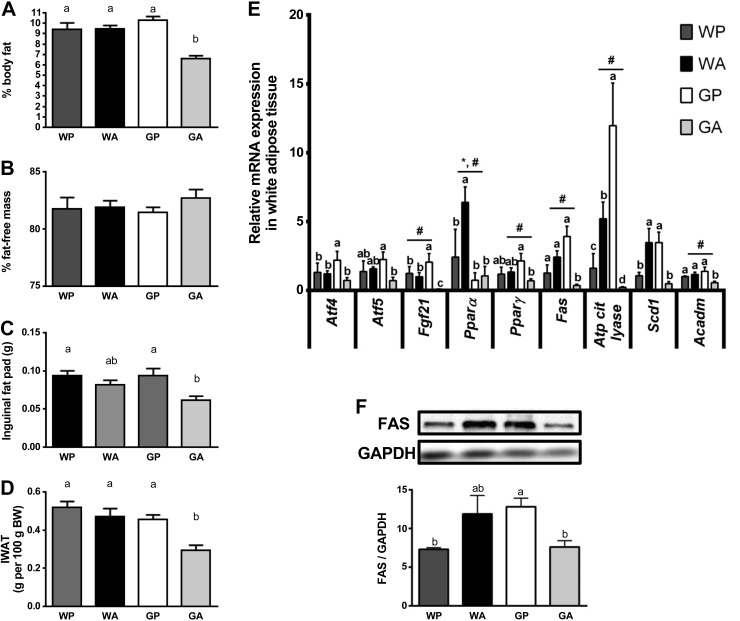
The antileukemic agent asparaginase triggers the amino acid response (AAR) in the liver by activating the eukaryotic initiation factor 2 (eIF2) kinase general control nonderepressible 2 (GCN2). To explore the mechanism by which AAR induction is necessary to mitigate hepatic lipid accumulation and prevent liver dysfunction during continued asparaginase treatment, wild-type and Gcn2 null mice were injected once daily with asparaginase or phosphate buffered saline for up to 14 days. Asparaginase induced mRNA expression of multiple AAR genes and greatly increased circulating concentrations of the metabolic hormone fibroblast growth factor 21 (FGF21) independent of food intake. Loss of Gcn2 precluded mRNA expression and circulating levels of FGF21 and blocked mRNA expression of multiple genes regulating lipid synthesis and metabolism including Fas, Ppara, Pparg, Acadm, and Scd1 in both liver and white adipose tissue. Furthermore, rates of triglyceride export and protein expression of apolipoproteinB-100 were significantly reduced in the livers of Gcn2 null mice treated with asparaginase, providing a mechanistic basis for the increase in hepatic lipid content. Loss of AAR-regulated antioxidant defenses in Gcn2 null livers was signified by reduced Gpx1 gene expression alongside increased lipid peroxidation. Substantial reductions in antithrombin III hepatic expression and activity in the blood of asparaginase-treated Gcn2 null mice indicated liver dysfunction. These results suggest that the ability of the liver to adapt to prolonged asparaginase treatment is influenced by GCN2-directed regulation of FGF21 and oxidative defenses, which, when lost, corresponds with maladaptive effects on lipid metabolism and hemostasis.
Keywords: adipose; amino acid response; antithrombin III; apolipoprotein B-100; eukaryotic initiation factor 2; fibroblast growth factor 21; general control nonderepressible 2; liver; mice; oxidative stress; steatosis.
Copyright © 2015 the American Physiological Society.
Figures
References
-
- Ai D, Baez JM, Jiang H, Conlon DM, Hernandez-Ono A, Frank-Kamenetsky M, Milstein S, Fitzgerald K, Murphy AJ, Woo CW, Strong A, Ginsberg HN, Tabas I, Rader DJ, Tall AR. Activation of ER stress and mTORC1 suppresses hepatic sortilin-1 levels in obese mice. J Clin Invest 122: 1677–1687, 2012. - PMC - PubMed
-
- Anthony TG, McDaniel BJ, Byerley RL, McGrath BC, Cavener DR, McNurlan MA, Wek RC. Preservation of liver protein synthesis during dietary leucine deprivation occurs at the expense of skeletal muscle mass in mice deleted for eIF2 kinase GCN2. J Biol Chem 279: 36553–36561, 2004. - PubMed
-
- Arriazu E, Perez de Obanos MP, Lopez-Zabalza MJ, Herraiz MT, Iraburu MJ. Amino acid deprivation decreases intracellular levels of reactive oxygen species in hepatic stellate cells. Cell Physiol Biochem 26: 281–290, 2010. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous