

Thrombospondin-1 restrains neutrophil granule serine protease function and regulates the innate immune response during Klebsiella pneumoniae infection
- PMID: 25492474
- PMCID: PMC4465063
- DOI: 10.1038/mi.2014.120
Thrombospondin-1 restrains neutrophil granule serine protease function and regulates the innate immune response during Klebsiella pneumoniae infection
Abstract
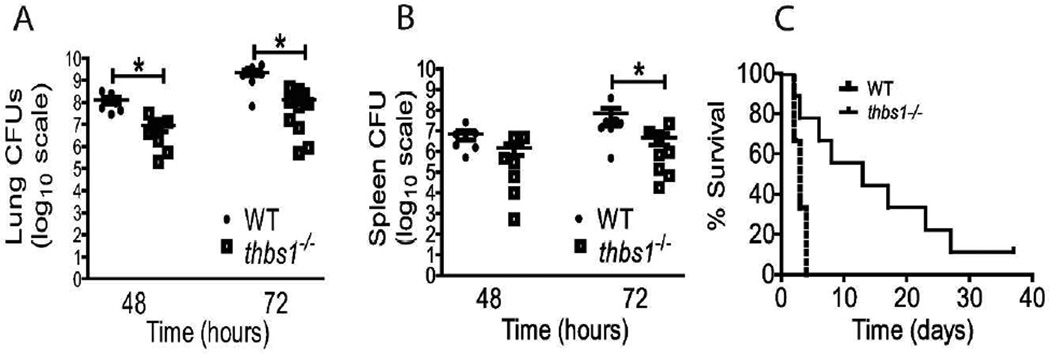
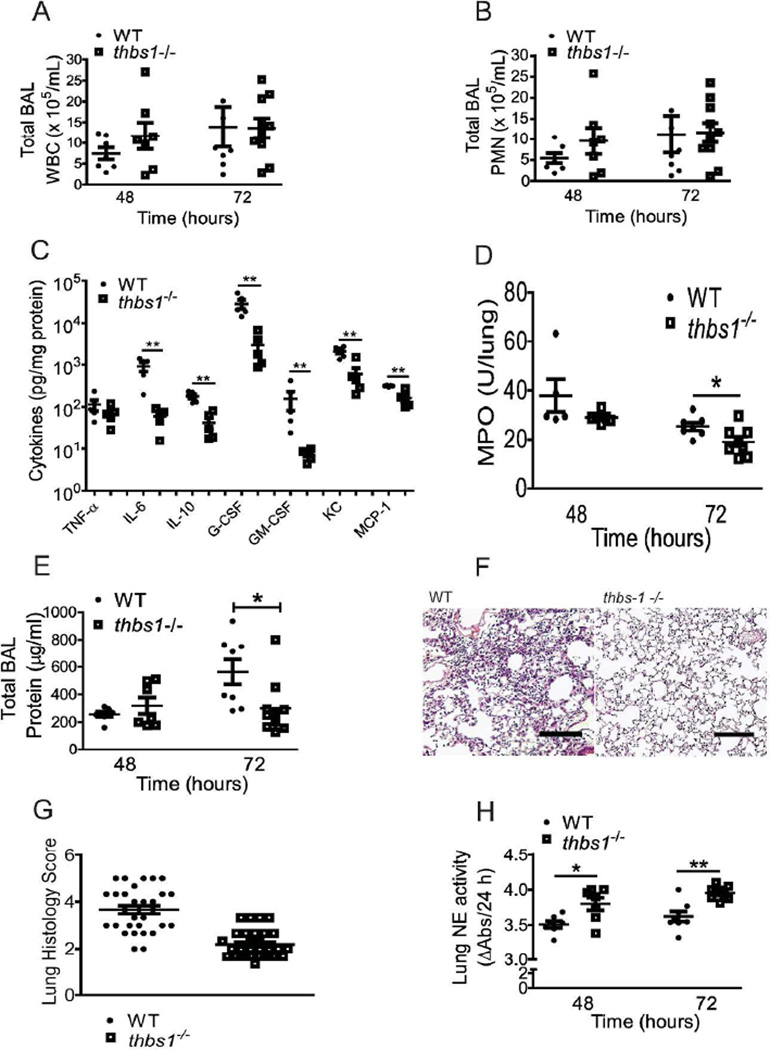
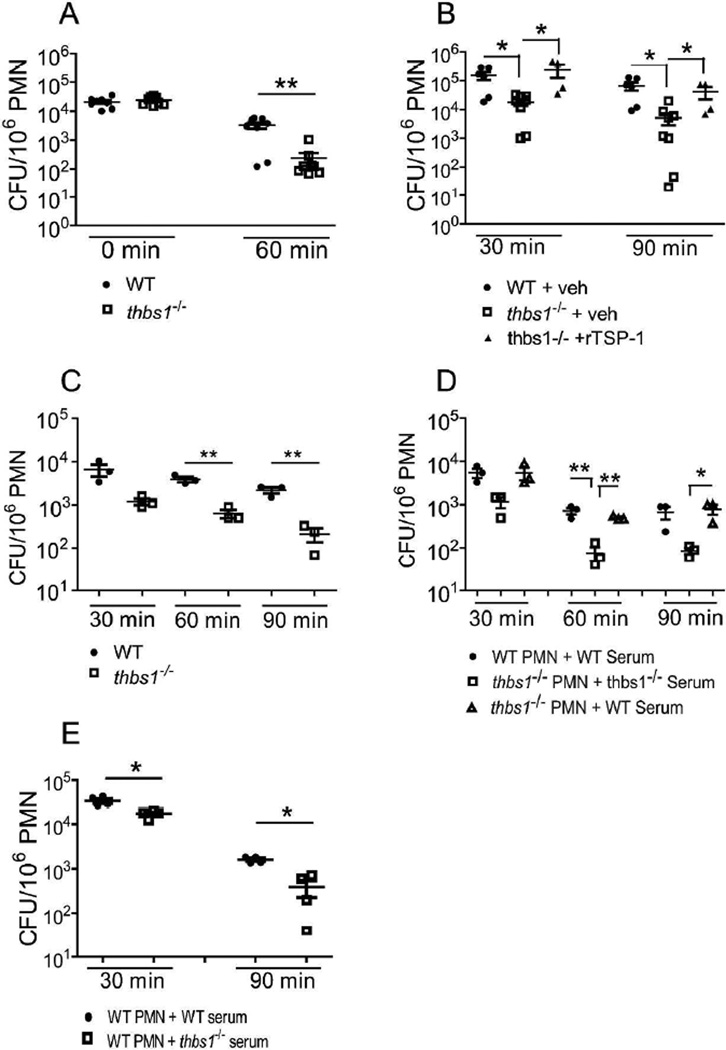
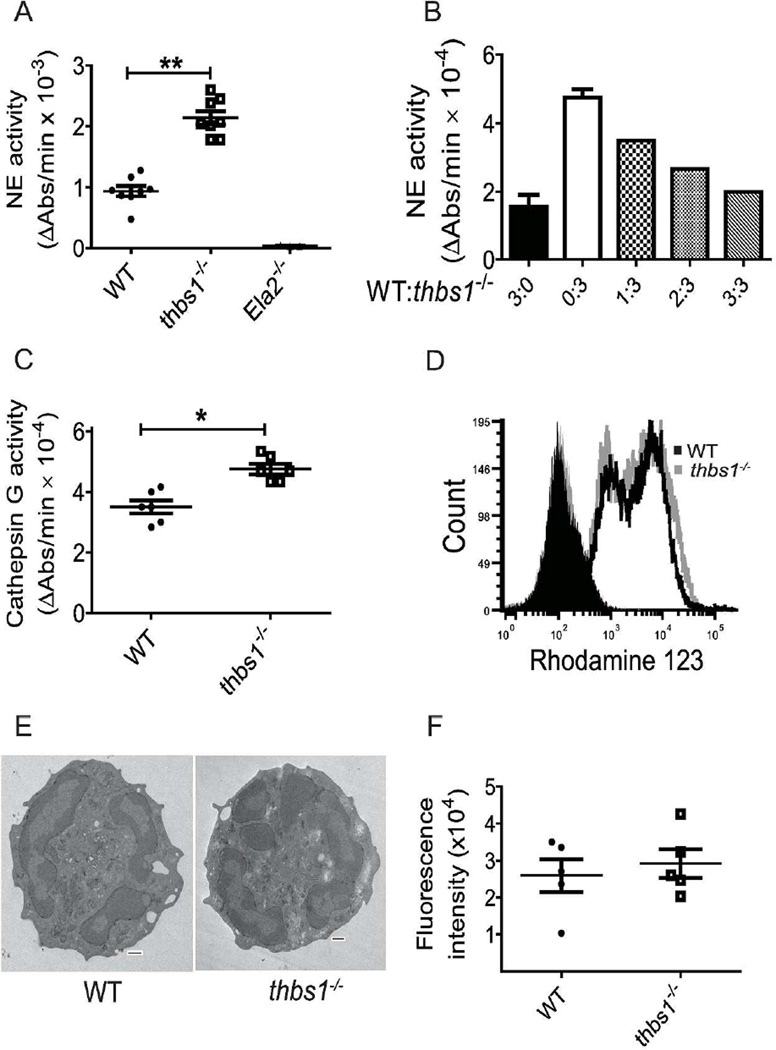
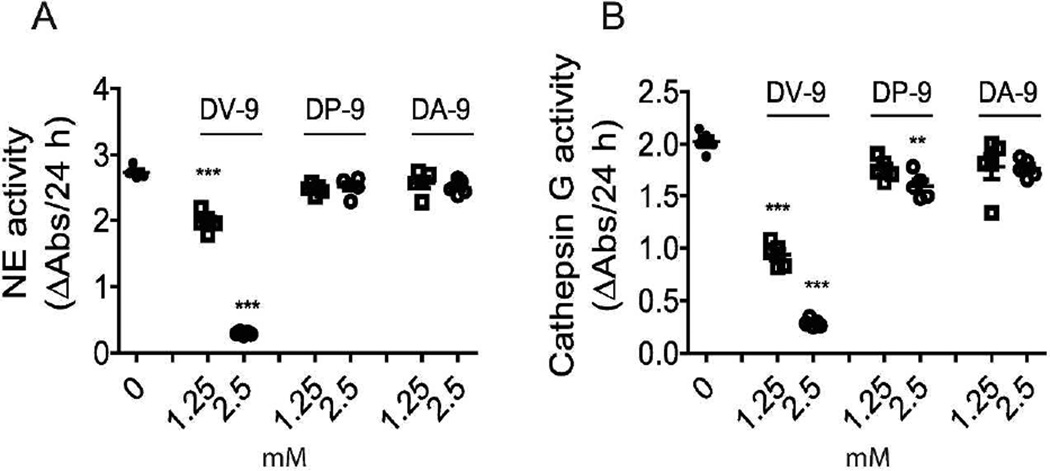
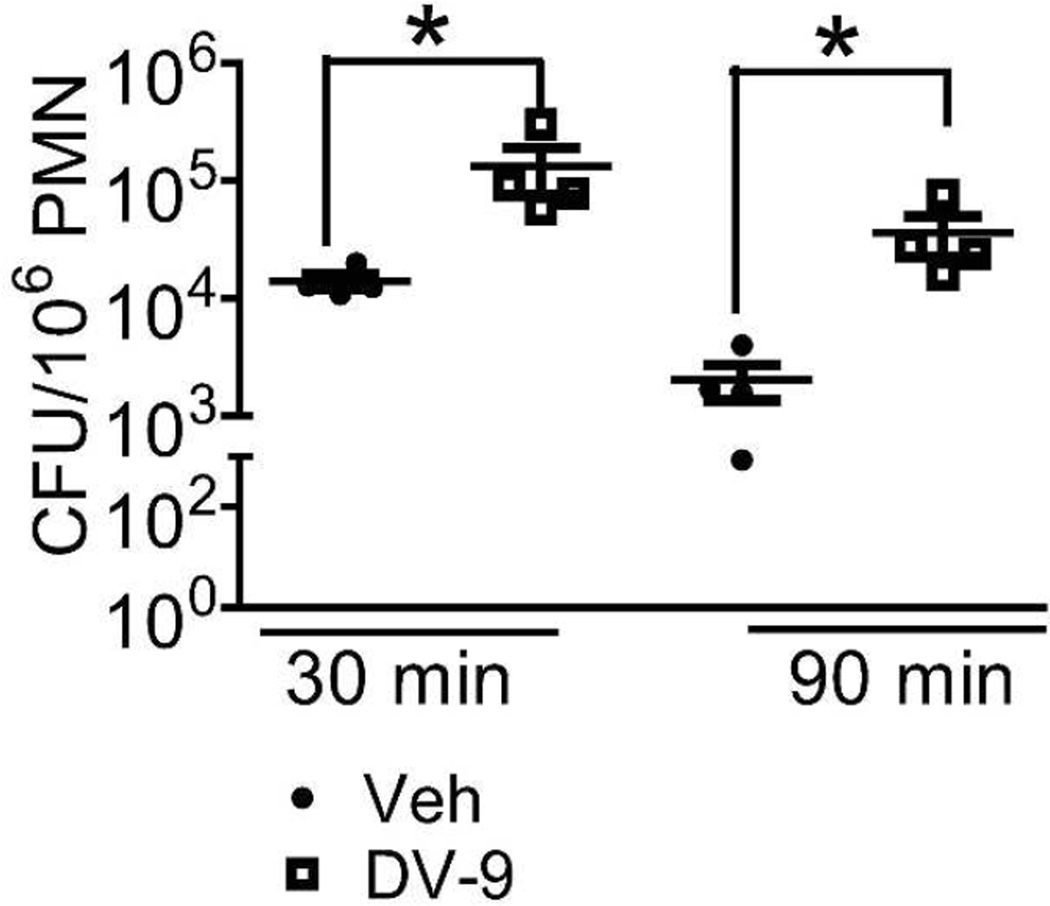
Neutrophil elastase (NE) and cathepsin G (CG) contribute to intracellular microbial killing but, if left unchecked and released extracellularly, promote tissue damage. Conversely, mechanisms that constrain neutrophil serine protease activity protect against tissue damage but may have the untoward effect of disabling the microbial killing arsenal. The host elaborates thrombospondin-1 (TSP-1), a matricellular protein released during inflammation, but its role during neutrophil activation following microbial pathogen challenge remains uncertain. Mice deficient in TSP-1 (thbs1(-/-)) showed enhanced lung bacterial clearance, reduced splenic dissemination, and increased survival compared with wild-type (WT) controls during intrapulmonary Klebsiella pneumoniae infection. More effective pathogen containment was associated with reduced burden of inflammation in thbs1(-/-) mouse lungs compared with WT controls. Lung NE activity was increased in thbs1(-/-) mice following K. pneumoniae challenge, and thbs1(-/-) neutrophils showed enhanced intracellular microbial killing that was abrogated with recombinant TSP-1 administration or WT serum. Thbs1(-/-) neutrophils exhibited enhanced NE and CG enzymatic activity, and a peptide corresponding to amino-acid residues 793-801 within the type-III repeat domain of TSP-1 bridled neutrophil proteolytic function and microbial killing in vitro. Thus, TSP-1 restrains proteolytic action during neutrophilic inflammation elicited by K. pneumoniae, providing a mechanism that may regulate the microbial killing arsenal.
Conflict of interest statement
Figures
References
-
- Hogg PJ, Owensby DA, Chesterman CN. Thrombospondin 1 is a tight-binding competitive inhibitor of neutrophil cathepsin G. Determination of the kinetic mechanism of inhibition and localization of cathepsin G binding to the thrombospondin 1 type 3 repeats. J Biol Chem. 1993;268:21811–21818. - PubMed
-
- Hogg PJ, Owensby DA, Mosher DF, Misenheimer TM, Chesterman CN. Thrombospondin is a tight-binding competitive inhibitor of neutrophil elastase. J Biol Chem. 1993;268:7139–7146. - PubMed
-
- Lawler J. The structural and functional properties of thrombospondin. Blood. 1986;67:1197–1209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
