

Triacylglycerol Storage in Lipid Droplets in Procyclic Trypanosoma brucei
- PMID: 25493940
- PMCID: PMC4262433
- DOI: 10.1371/journal.pone.0114628
Triacylglycerol Storage in Lipid Droplets in Procyclic Trypanosoma brucei
Abstract
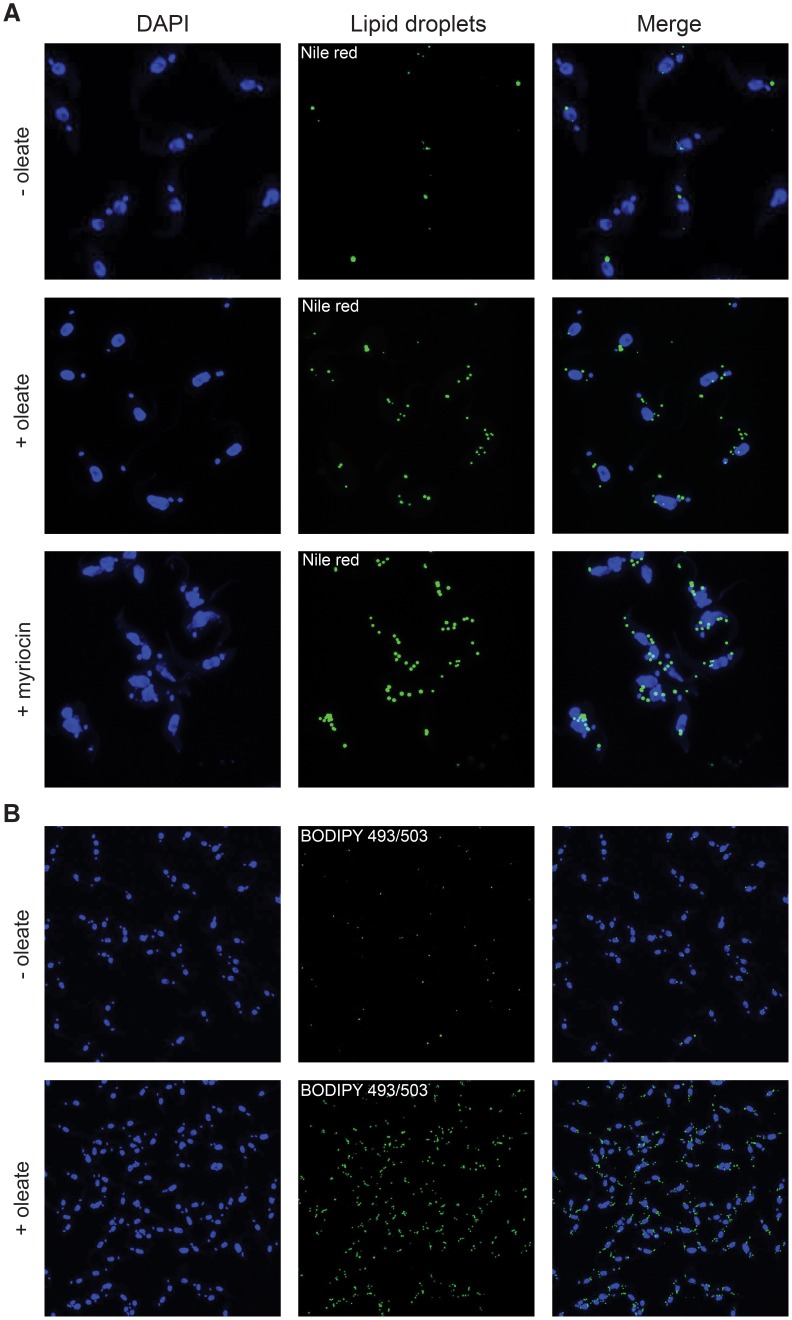
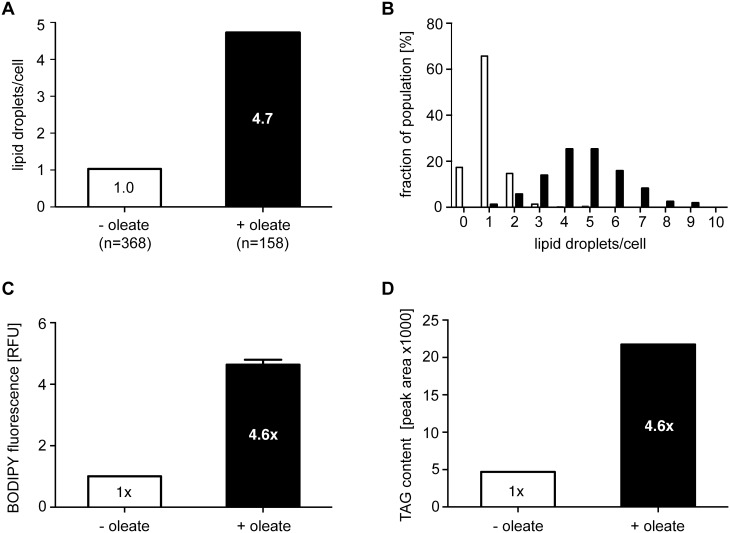
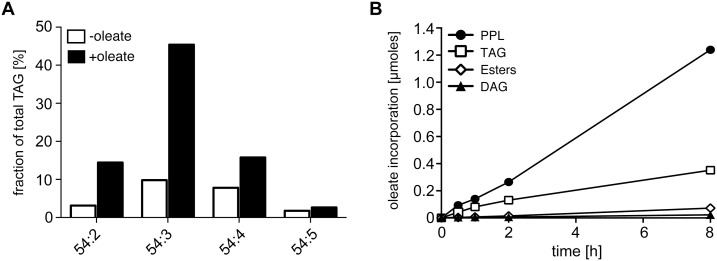
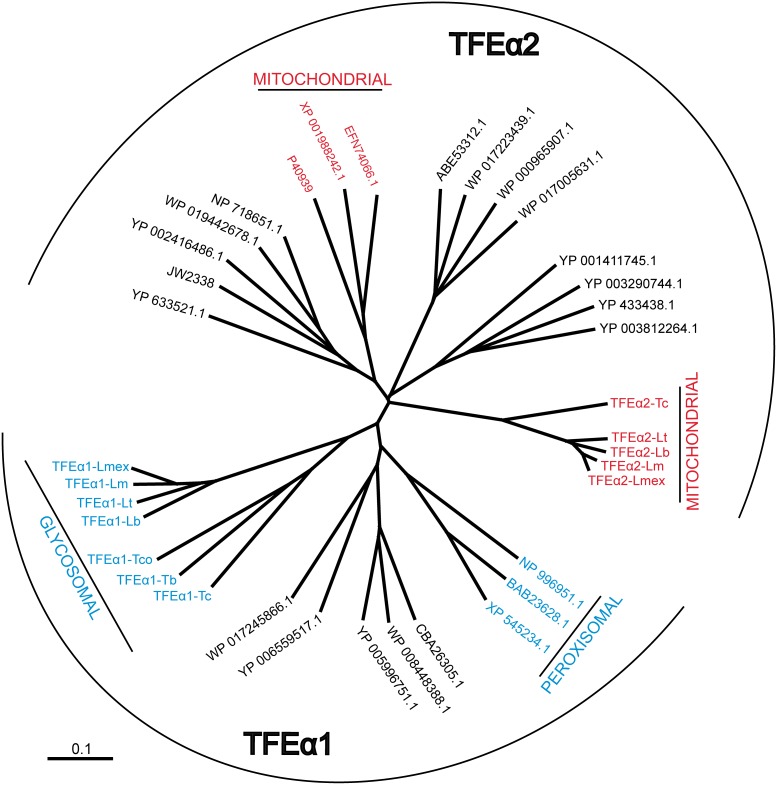
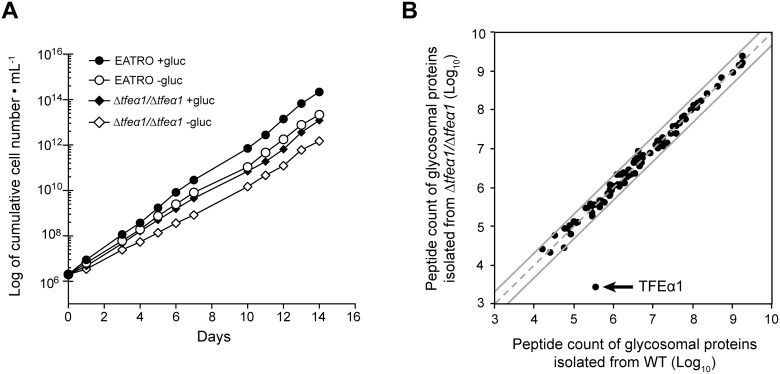
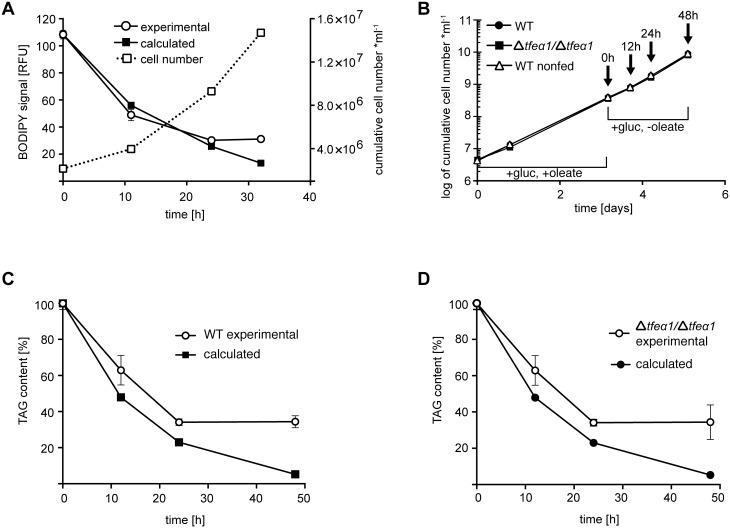
Carbon storage is likely to enable adaptation of trypanosomes to nutritional challenges or bottlenecks during their stage development and migration in the tsetse. Lipid droplets are candidates for this function. This report shows that feeding of T. brucei with oleate results in a 4-5 fold increase in the number of lipid droplets, as quantified by confocal fluorescence microscopy and by flow cytometry of BODIPY 493/503-stained cells. The triacylglycerol (TAG) content also increased 4-5 fold, and labeled oleate is incorporated into TAG. Fatty acid carbon can thus be stored as TAG in lipid droplets under physiological growth conditions in procyclic T. brucei. β-oxidation has been suggested as a possible catabolic pathway for lipids in T. brucei. A single candidate gene, TFEα1 with coding capacity for a subunit of the trifunctional enzyme complex was identified. TFEα1 is expressed in procyclic T. brucei and present in glycosomal proteomes, Unexpectedly, a TFEα1 gene knock-out mutant still expressed wild-type levels of previously reported NADP-dependent 3-hydroxyacyl-CoA dehydrogenase activity, and therefore, another gene encodes this enzymatic activity. Homozygous Δtfeα1/Δtfeα1 null mutant cells show a normal growth rate and an unchanged glycosomal proteome in procyclic T. brucei. The decay kinetics of accumulated lipid droplets upon oleate withdrawal can be fully accounted for by the dilution effect of cell division in wild-type and Δtfeα1/Δtfeα1 cells. The absence of net catabolism of stored TAG in procyclic T. brucei, even under strictly glucose-free conditions, does not formally exclude a flux through TAG, in which biosynthesis equals catabolism. Also, the possibility remains that TAG catabolism is completely repressed by other carbon sources in culture media or developmentally activated in post-procyclic stages in the tsetse.
Conflict of interest statement
Figures
References
-
- Martin S, Parton RG (2006) Lipid droplets: a unified view of a dynamic organelle. Nat Rev Mol Cell Biol 7:373–378. - PubMed
-
- Igal RA, Coleman RA (1996) Acylglycerol recycling from triacylglycerol to phospholipid, not lipase activity, is defective in neutral lipid storage disease fibroblasts. J Biol Chem 271:16644–16651. - PubMed
-
- Igal RA, Coleman RA (1998) Neutral lipid storage disease: a genetic disorder with abnormalities in the regulation of phospholipid metabolism. J Lipid Res 39:31–43. - PubMed
-
- Leber R, Zinser E, Hrastnik C, Paltauf F, Daum G (1995) Export of steryl esters from lipid particles and release of free sterols in the yeast, Saccharomyces cerevisiae. Biochim Biophys Acta 1234:119–126. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
