

The effect of neurospecific knockdown of candidate genes for locomotor behavior and sound production in Drosophila melanogaster
- PMID: 25494872
- PMCID: PMC4594543
- DOI: 10.4161/19336934.2014.983389
The effect of neurospecific knockdown of candidate genes for locomotor behavior and sound production in Drosophila melanogaster
Abstract
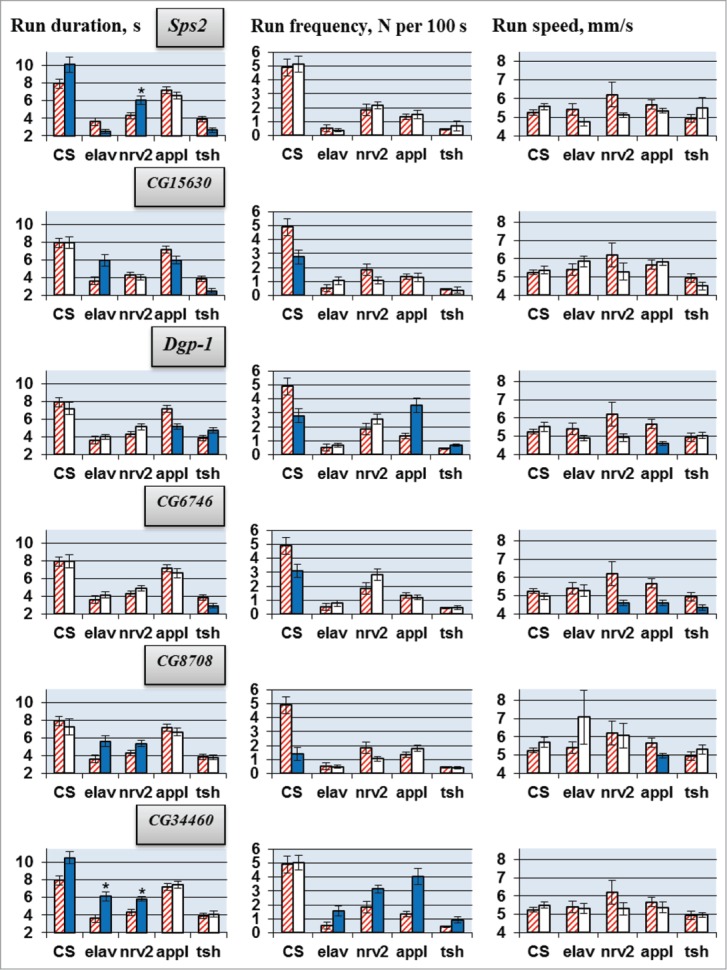
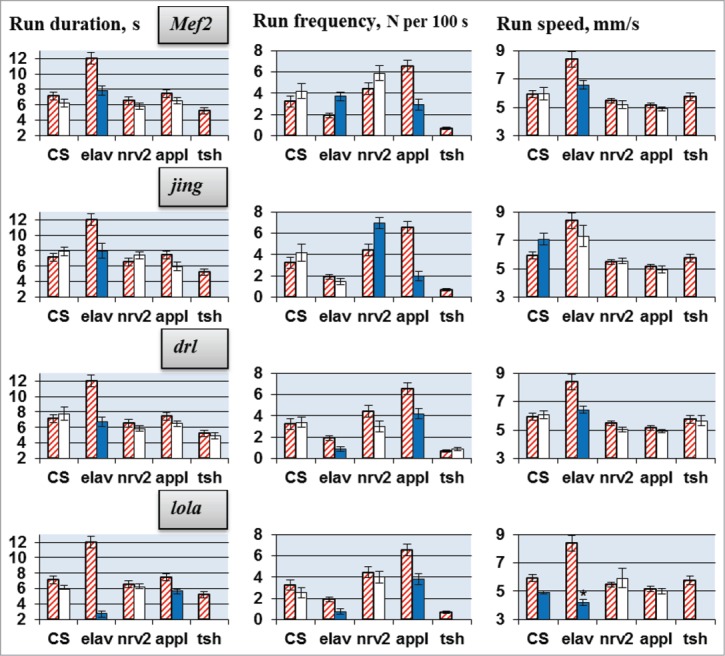
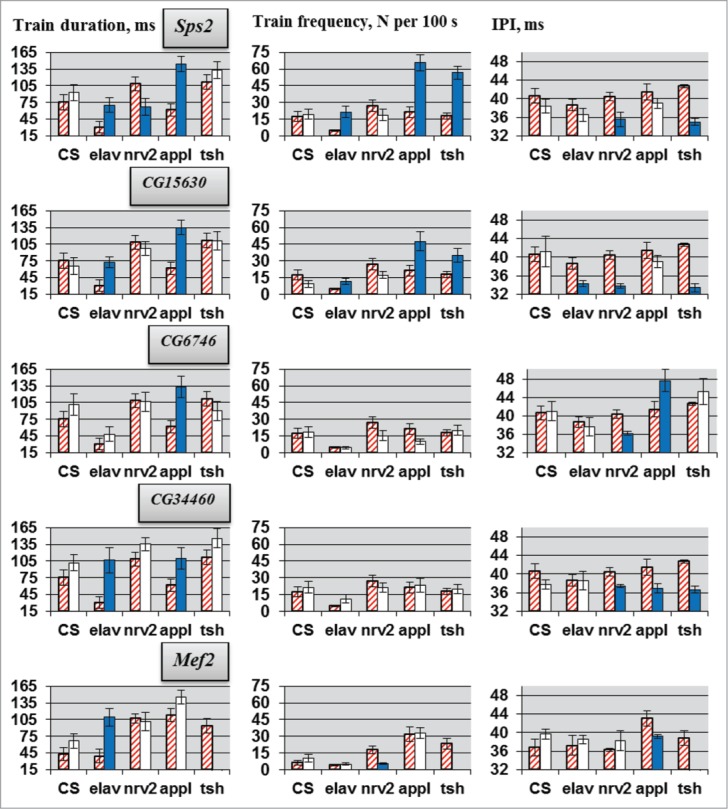
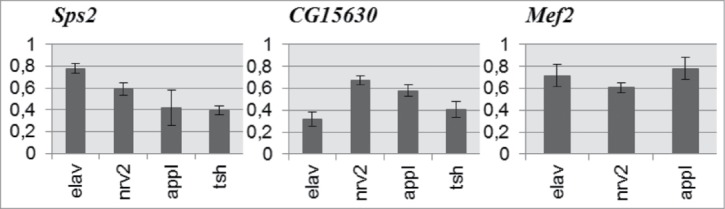
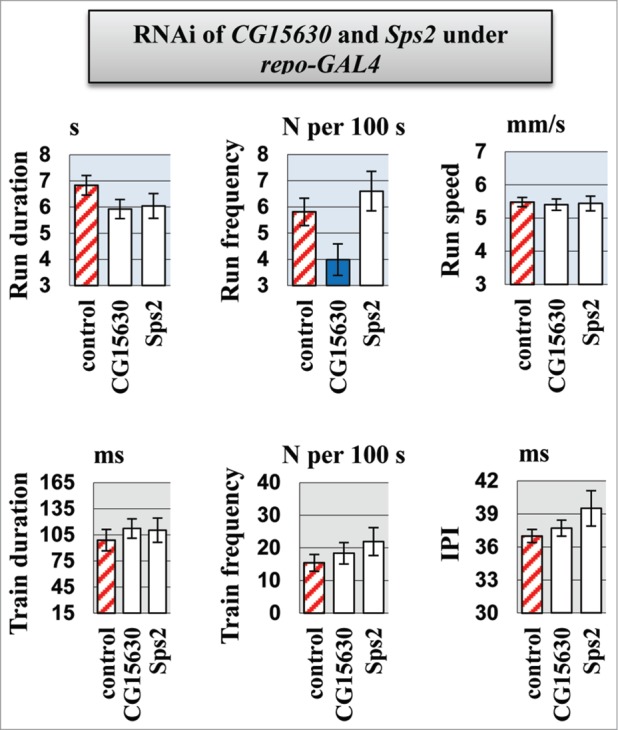
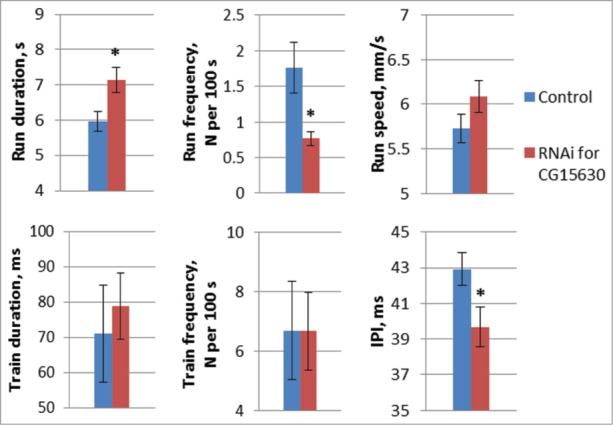
Molecular mechanisms underlying the functioning of central pattern generators (CPGs) are poorly understood. Investigations using genetic approaches in the model organism Drosophila may help to identify unknown molecular players participating in the formation or control of motor patterns. Here we report Drosophila genes as candidates for involvement in the neural mechanisms responsible for motor functions, such as locomotion and courtship song. Twenty-two Drosophila lines, used for gene identification, were isolated from a previously created collection of 1064 lines, each carrying a P element insertion in one of the autosomes. The lines displayed extreme deviations in locomotor and/or courtship song parameters compared with the whole collection. The behavioral consequences of CNS-specific RNAi-mediated knockdowns for 10 identified genes were estimated. The most prominent changes in the courtship song interpulse interval (IPI) were seen in flies with Sps2 or CG15630 knockdown. Glia-specific knockdown of these genes produced no effect on the IPI. Estrogen-induced knockdown of CG15630 in adults reduced the IPI. The product of the CNS-specific gene, CG15630 (a predicted cell surface receptor), is likely to be directly involved in the functioning of the CPG generating the pulse song pattern. Future studies should ascertain its functional role in the neurons that constitute the song CPG. Other genes (Sps2, CG34460), whose CNS-specific knockdown resulted in IPI reduction, are also worthy of detailed examination.
Keywords: BDSC; Bloomington Drosophila Stock Center; CNS; Canton-S strain; FITC; Drosophila; P element insertion; Ponce de Leon P element construct; RNAi; RNA interference; RNA interference; shRNA; Transgenic RNAi Project; UAS; Vienna Drosophila RNAi Center; candidate genes; central nervous system; CPG; central pattern generator; CS; central pattern generators, courtship song; fluorescein isothiocyanate; Gal4; green fluorescent protein; IPI; interpulse interval; LA; locomotion; locomotor activity; PBS; phosphate-buffered saline; PCR; polymerase chain reaction; PdL; small hairpin RNA; TRiP; upstream activation sequence; VDRC; yeast transcription activator protein Gal4; GFP.
Figures
References
-
- Briggman KL, Kristan WB. Multifunctional pattern-generating circuits. Annu Rev Neurosci 2008; 31:271-94;PMID:18558856; http://dx.doi.org/ 10.1146/annurev.neuro.31.060407.125552 - DOI - PubMed
-
- Marder E, Bucher D. Understanding circuit dynamics using the stomatogastric nervous system of lobsters and crabs. Annu Rev Physiol 2007; 69:291-316; PMID:17009928; http://dx.doi.org/ 10.1146/annurev.physiol.69.031905.161516 - DOI - PubMed
-
- Kiehn O. Development and functional organization of spinal locomotor circuits. Curr Opin Neurobiol 2011; 21:100-9; PMID:20889331; http://dx.doi.org/ 10.1016/j.conb.2010.09.004 - DOI - PubMed
-
- Lu TZ, Feng Z-P. NALCN: A regulator of pacemaker activity. Mol Neurobiol 2012; 45:415-23; PMID:22476981; http://dx.doi.org/ 10.1007/s12035-012-8260-2 - DOI - PubMed
-
- Nash HA, Scott RL, Lear BC, Allada R. An unusual cation channel mediates photic control of locomotion in Drosophila. Curr Biol 2002; 12:2152-8; PMID:12498692; http://dx.doi.org/ 10.1016/S0960-9822(02)01358-1 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases