

Catalytic surface radical in dye-decolorizing peroxidase: a computational, spectroscopic and site-directed mutagenesis study
- PMID: 25495127
- PMCID: PMC4357238
- DOI: 10.1042/BJ20141211
Catalytic surface radical in dye-decolorizing peroxidase: a computational, spectroscopic and site-directed mutagenesis study
Abstract
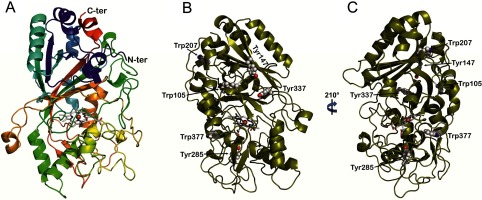
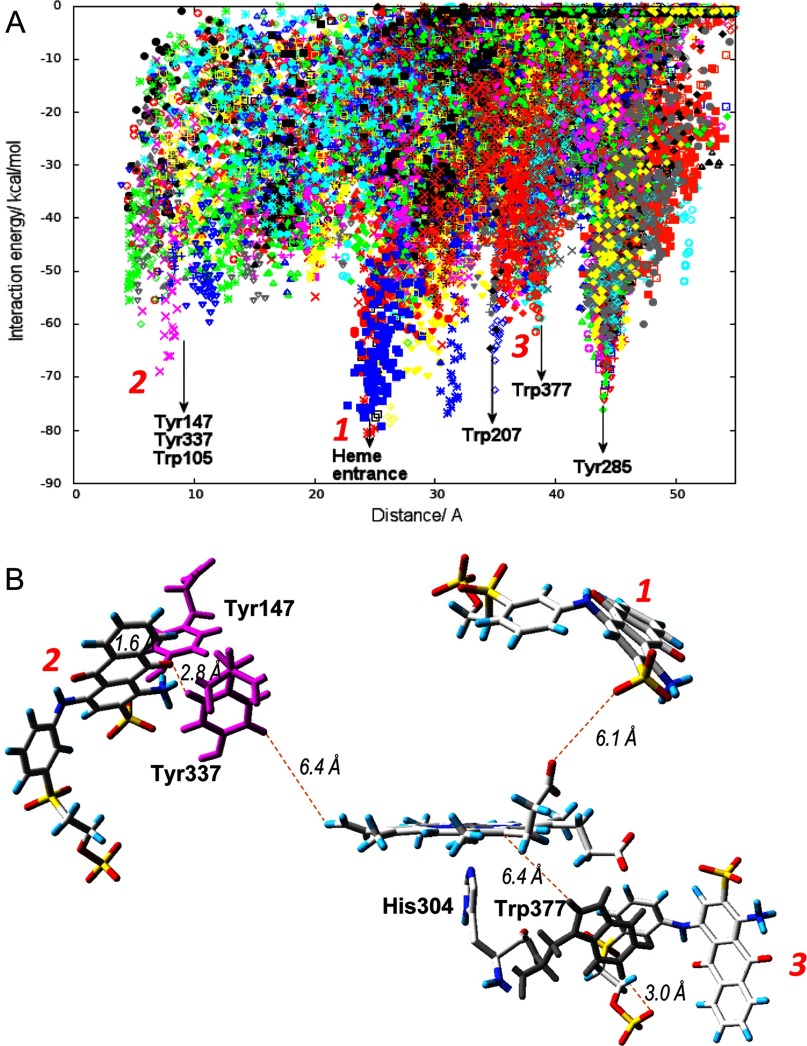
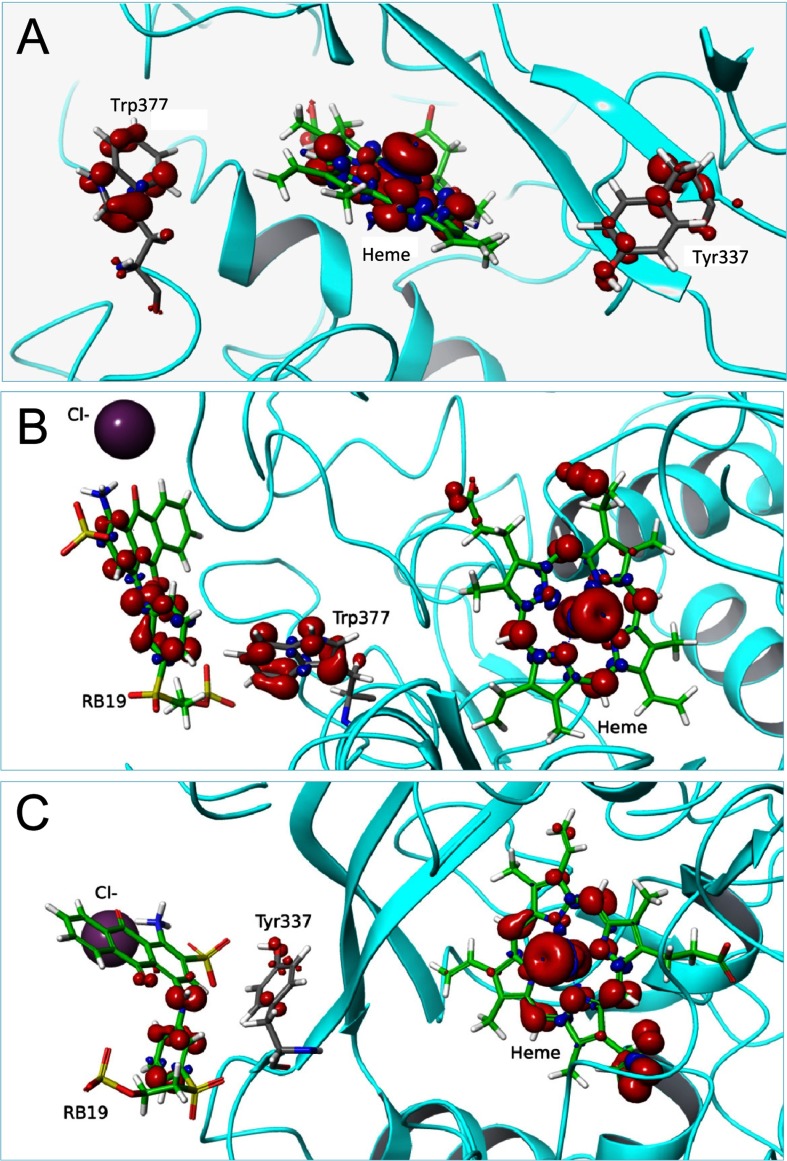
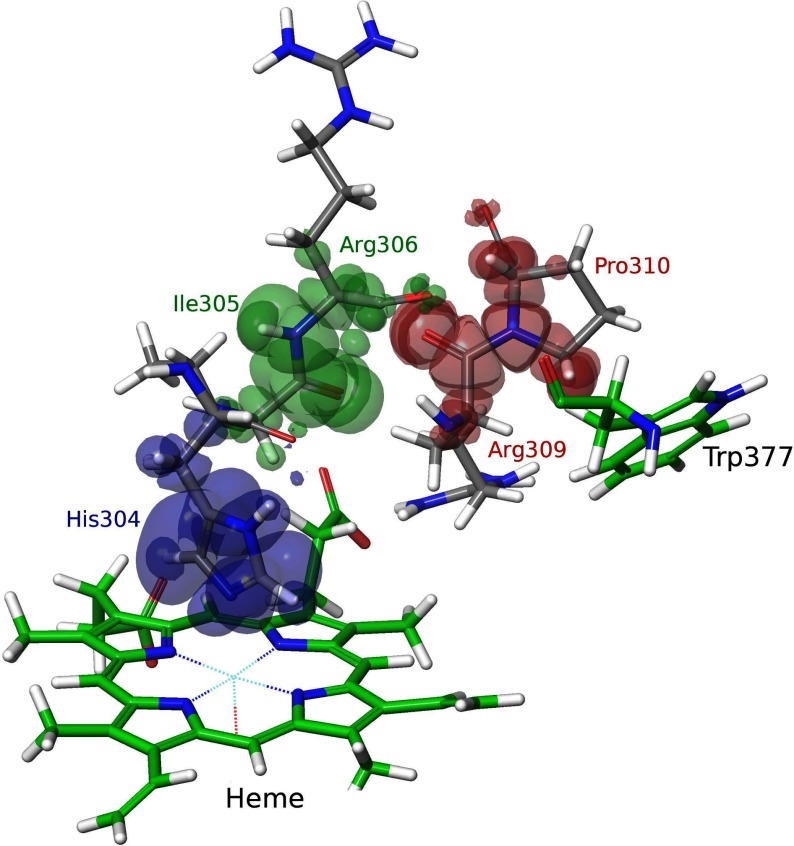
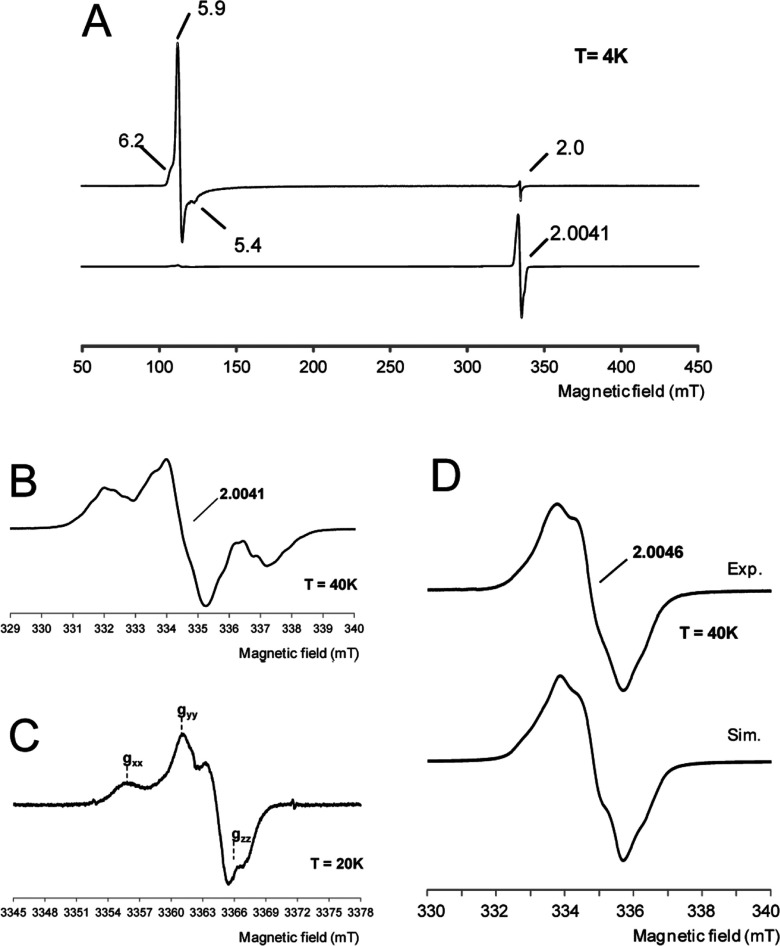
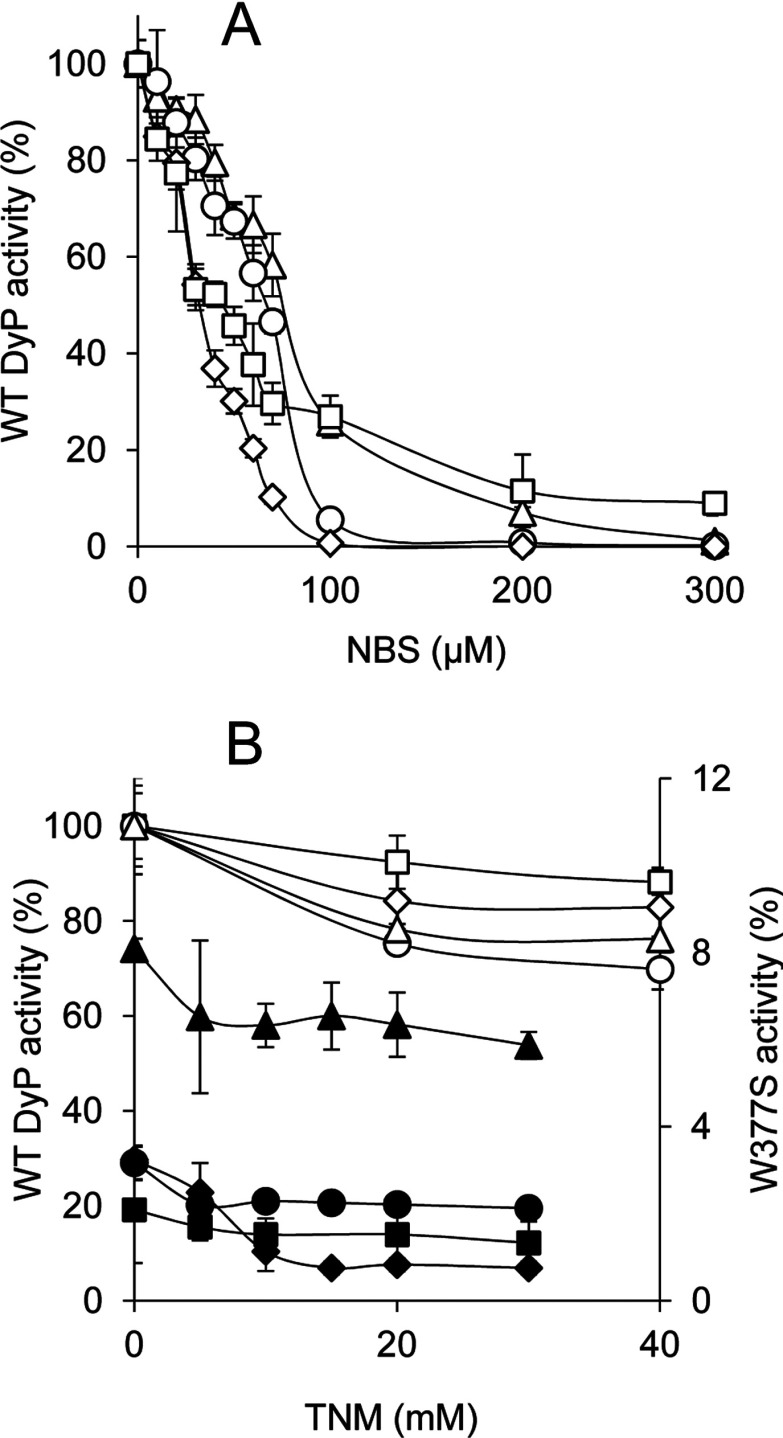
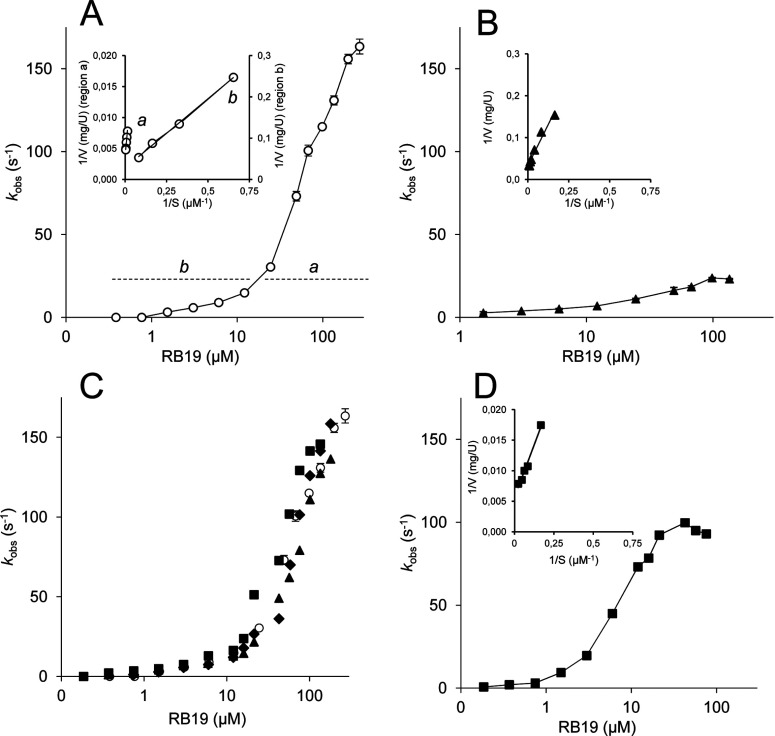
Dye-decolorizing peroxidase (DyP) of Auricularia auricula-judae has been expressed in Escherichia coli as a representative of a new DyP family, and subjected to mutagenic, spectroscopic, crystallographic and computational studies. The crystal structure of DyP shows a buried haem cofactor, and surface tryptophan and tyrosine residues potentially involved in long-range electron transfer from bulky dyes. Simulations using PELE (Protein Energy Landscape Exploration) software provided several binding-energy optima for the anthraquinone-type RB19 (Reactive Blue 19) near the above aromatic residues and the haem access-channel. Subsequent QM/MM (quantum mechanics/molecular mechanics) calculations showed a higher tendency of Trp-377 than other exposed haem-neighbouring residues to harbour a catalytic protein radical, and identified the electron-transfer pathway. The existence of such a radical in H₂O₂-activated DyP was shown by low-temperature EPR, being identified as a mixed tryptophanyl/tyrosyl radical in multifrequency experiments. The signal was dominated by the Trp-377 neutral radical contribution, which disappeared in the W377S variant, and included a tyrosyl contribution assigned to Tyr-337 after analysing the W377S spectra. Kinetics of substrate oxidation by DyP suggests the existence of high- and low-turnover sites. The high-turnover site for oxidation of RB19 (k(cat) > 200 s⁻¹) and other DyP substrates was assigned to Trp-377 since it was absent from the W377S variant. The low-turnover site/s (RB19 k(cat) ~20 s⁻¹) could correspond to the haem access-channel, since activity was decreased when the haem channel was occluded by the G169L mutation. If a tyrosine residue is also involved, it will be different from Tyr-337 since all activities are largely unaffected in the Y337S variant.
Figures
References
-
- Ruiz-Dueñas F.J., Martínez A.T. Structural and functional features of peroxidases with a potential as industrial biocatalysts. In: Torres E., Ayala M., editors. Biocatalysts Based on Heme Peroxidases. Berlin: Springer-Verlag; 2010. pp. 37–59. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous