

Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes
- PMID: 25495187
- PMCID: PMC4407914
- DOI: 10.1111/jeb.12570
Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes
Abstract
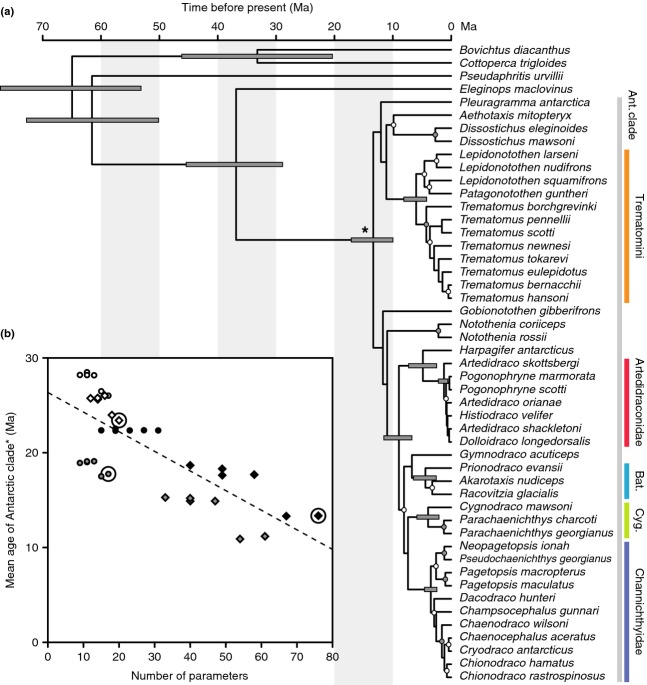
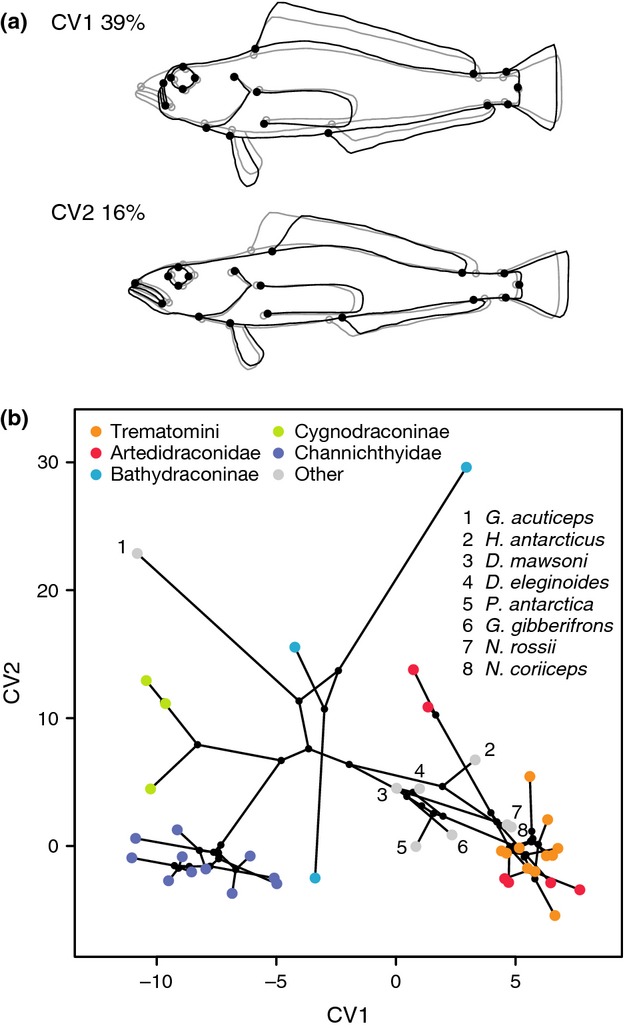
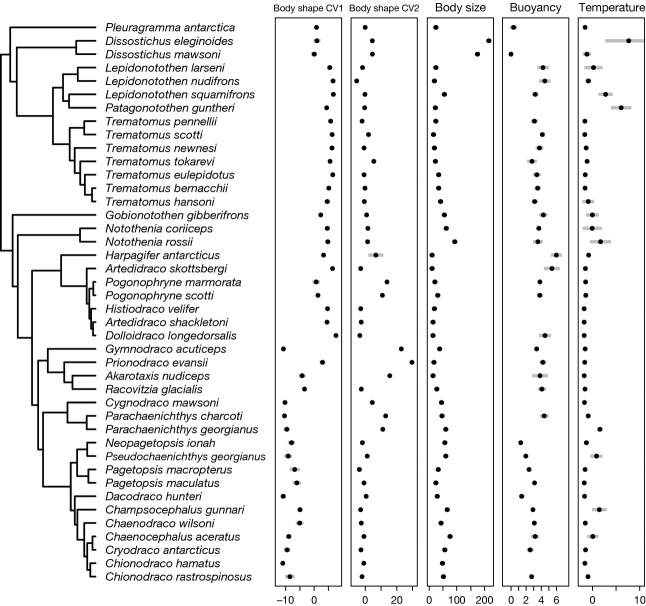
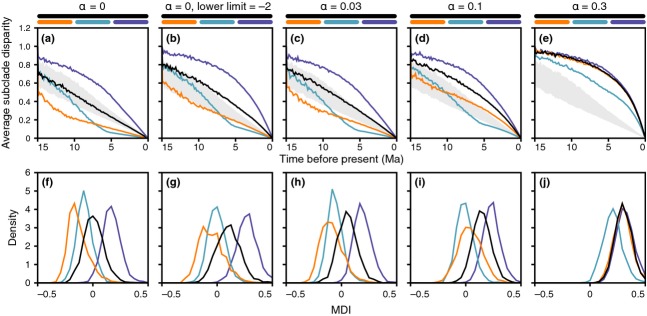
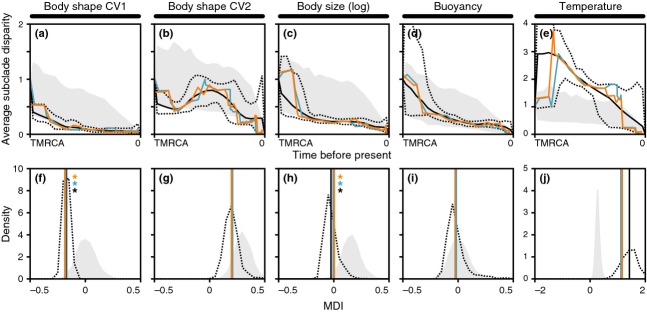
According to theory, adaptive radiation is triggered by ecological opportunity that can arise through the colonization of new habitats, the extinction of antagonists or the origin of key innovations. In the course of an adaptive radiation, diversification and morphological evolution are expected to slow down after an initial phase of rapid adaptation to vacant ecological niches, followed by speciation. Such 'early bursts' of diversification are thought to occur because niche space becomes increasingly filled over time. The diversification of Antarctic notothenioid fishes into over 120 species has become one of the prime examples of adaptive radiation in the marine realm and has likely been triggered by an evolutionary key innovation in the form of the emergence of antifreeze glycoproteins. Here, we test, using a novel time-calibrated phylogeny of 49 species and five traits that characterize notothenioid body size and shape as well as buoyancy adaptations and habitat preferences, whether the notothenioid adaptive radiation is compatible with an early burst scenario. Extensive Bayesian model comparison shows that phylogenetic age estimates are highly dependent on model choice and that models with unlinked gene trees are generally better supported and result in younger age estimates. We find strong evidence for elevated diversification rates in Antarctic notothenioids compared to outgroups, yet no sign of rate heterogeneity in the course of the radiation, except that the notothenioid family Artedidraconidae appears to show secondarily elevated diversification rates. We further observe an early burst in trophic morphology, suggesting that the notothenioid radiation proceeds in stages similar to other prominent examples of adaptive radiation.
Keywords: adaptive radiation; early burst; geometric morphometrics; incomplete lineage sorting; species tree.
© 2014 The Authors. Journal of Evolutionary Biology published by John Wiley & Sons Ltd on behalf of European Society for Evolutionary Biology.
Figures
References
-
- Ackerly DD, Schwilk DW. Webb CO. Niche evolution and adaptive radiation: testing the order of trait divergence. Ecology. 2006;87:S50–S61. - PubMed
-
- Adams CE. Huntingford FA. The functional significance of inherited differences in feeding morphology in a sympatric polymorphic population of Arctic charr. Evol. Ecol. 2002;16:15–25.
-
- Balushkin AV. Morphology, classification, and evolution of Notothenioid fishes of the Southern Ocean (Notothenioidei, Perciformes) J. Ichthyol. 2000;40:S74–S109.
-
- Bargelloni L, Ritchie PA, Patarnello T, Battaglia B, Lambert DM. Meyer A. Molecular evolution at subzero temperatures: mitochondrial and nuclear phylogenies of fishes from Antarctica (suborder Notothenioidei), and the evolution of antifreeze glycopeptides. Mol. Biol. Evol. 1994;11:854–863. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
