

Molecular mechanisms of the microsomal mixed function oxidases and biological and pathological implications
- PMID: 25498968
- PMCID: PMC4309856
- DOI: 10.1016/j.redox.2014.11.008
Molecular mechanisms of the microsomal mixed function oxidases and biological and pathological implications
Abstract
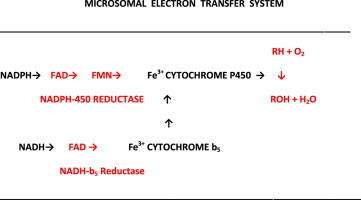
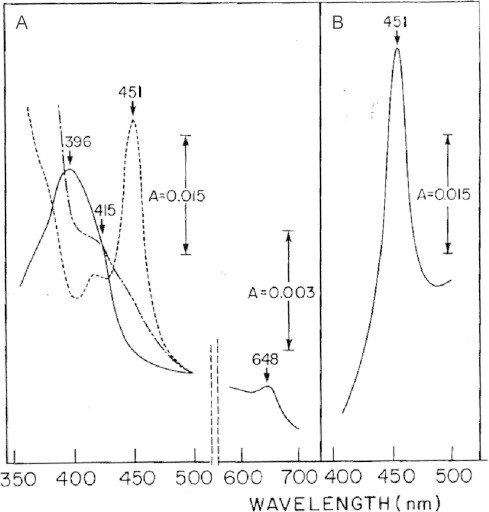
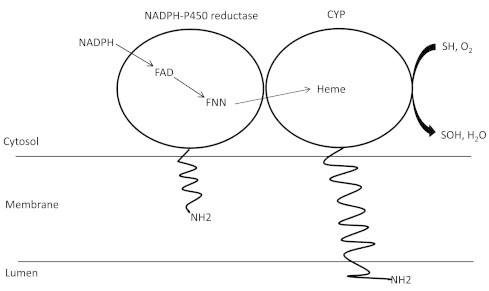
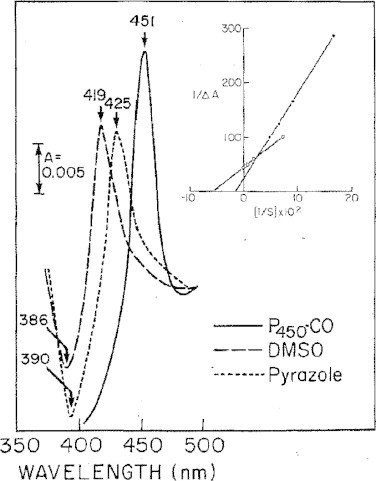
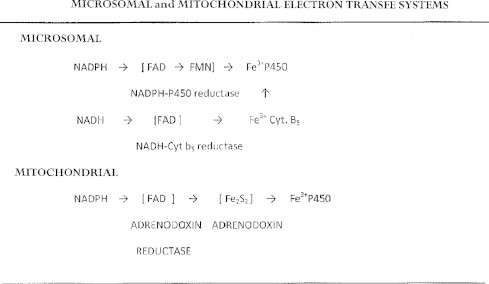
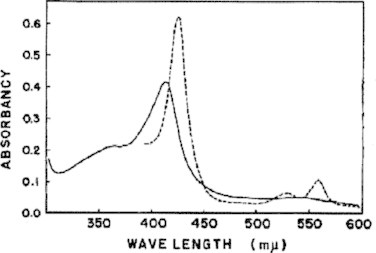
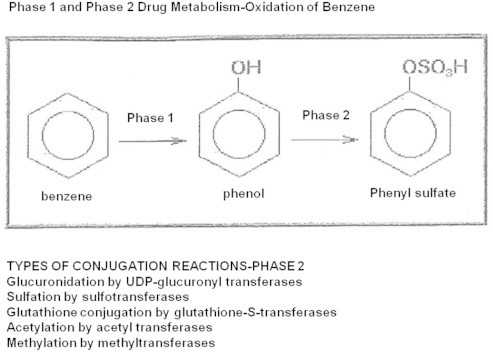
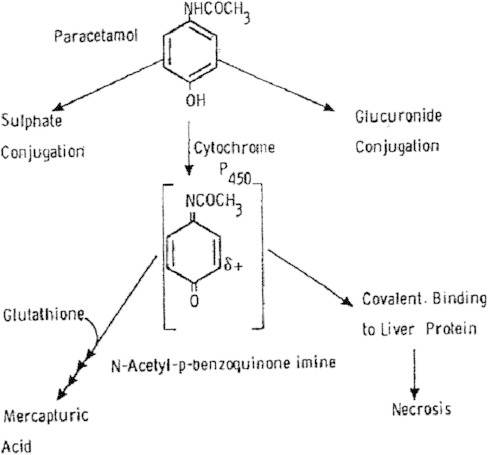
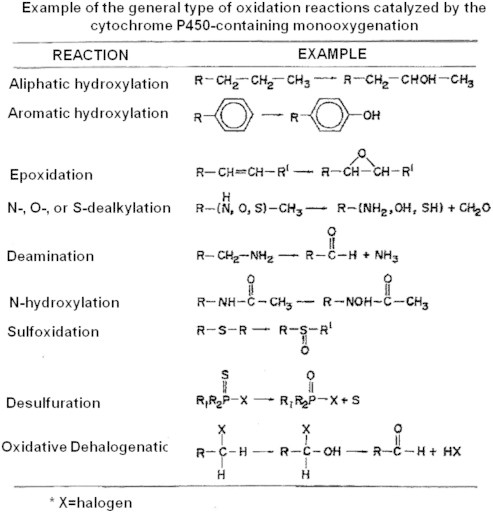
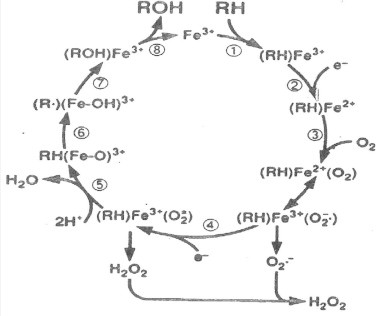
The cytochrome P450 mixed function oxidase enzymes play a major role in the metabolism of important endogenous substrates as well as in the biotransformation of xenobiotics. The liver P450 system is the most active in metabolism of exogenous substrates. This review briefly describes the liver P450 (CYP) mixed function oxidase system with respect to its enzymatic components and functions. Electron transfer by the NADPH-P450 oxidoreductase is required for reduction of the heme of P450, necessary for binding of molecular oxygen. Binding of substrates to P450 produce substrate binding spectra. The P450 catalytic cycle is complex and rate-limiting steps are not clear. Many types of chemical reactions can be catalyzed by P450 enzymes, making this family among the most diverse catalysts known. There are multiple forms of P450s arranged into families based on structural homology. The major drug metabolizing CYPs are discussed with respect to typical substrates, inducers and inhibitors and their polymorphic forms. The composition of CYPs in humans varies considerably among individuals because of sex and age differences, the influence of diet, liver disease, presence of potential inducers and/or inhibitors. Because of such factors and CYP polymorphisms, and overlapping drug specificity, there is a large variability in the content and composition of P450 enzymes among individuals. This can result in large variations in drug metabolism by humans and often can contribute to drug-drug interactions and adverse drug reactions. Because of many of the above factors, especially CYP polymorphisms, there has been much interest in personalized medicine especially with respect to which CYPs and which of their polymorphic forms are present in order to attempt to determine what drug therapy and what dosage would reflect the best therapeutic strategy in treating individual patients.
Keywords: Cytochrome P450; Liver microsomal drug metabolism system; NADPH-cytochrome P450 reductase; P450 catalytic cycle; P450 induction; P450 multiple forms; P450 polymorphisms; Personalized medicine.
Copyright © 2014 The Author. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Nebert D.W., Nelson D.R., Coon M.J., Estabrook R.W., Feyereisen R., Fujii-Kuriyama Y., Gonzalez F.J., Guengerich F.P., Gunsalus I.C., Johnson E.F., Loper J.C., Sato R., Waterman M.R., Waxman D.J. The P450 superfamily: update on new sequences, gene mapping, and recommended nomenclature. DNA and Cell Biology. 1991;10:1–4. - PubMed
-
- McKinnon R.A., Sorich M.J., Ward M.B. Cytochrome P450 Part 1: multiplicity and function. Journal of Research in Pharmacy Practice. 2008;38:55–57.
-
- Garfinkel D. Studies on pig liver microsomes. I. Enzymic and pigment composition of different microsomal fractions. Archives of Biochemistry and Biophysics. 1958;77:493–509. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
