

Genome-wide analysis of leafbladeless1-regulated and phased small RNAs underscores the importance of the TAS3 ta-siRNA pathway to maize development
- PMID: 25503246
- PMCID: PMC4263373
- DOI: 10.1371/journal.pgen.1004826
Genome-wide analysis of leafbladeless1-regulated and phased small RNAs underscores the importance of the TAS3 ta-siRNA pathway to maize development
Abstract
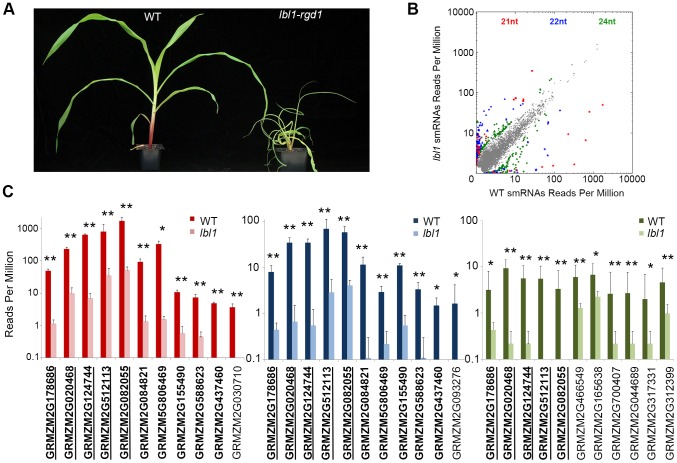
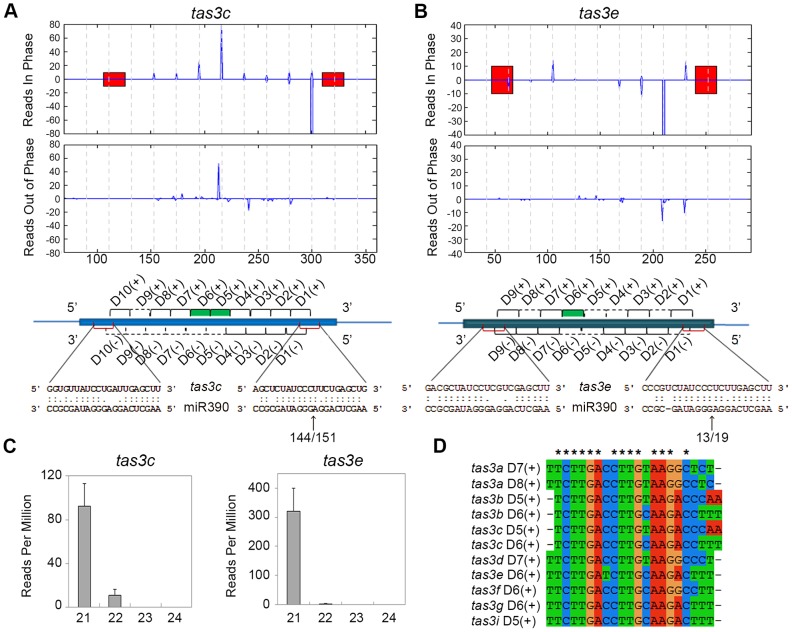
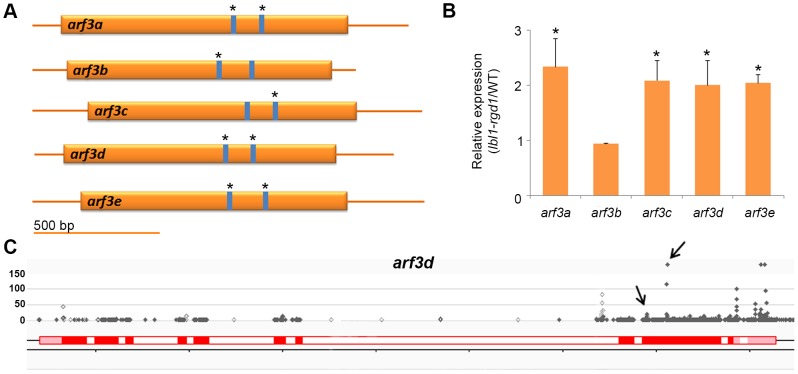
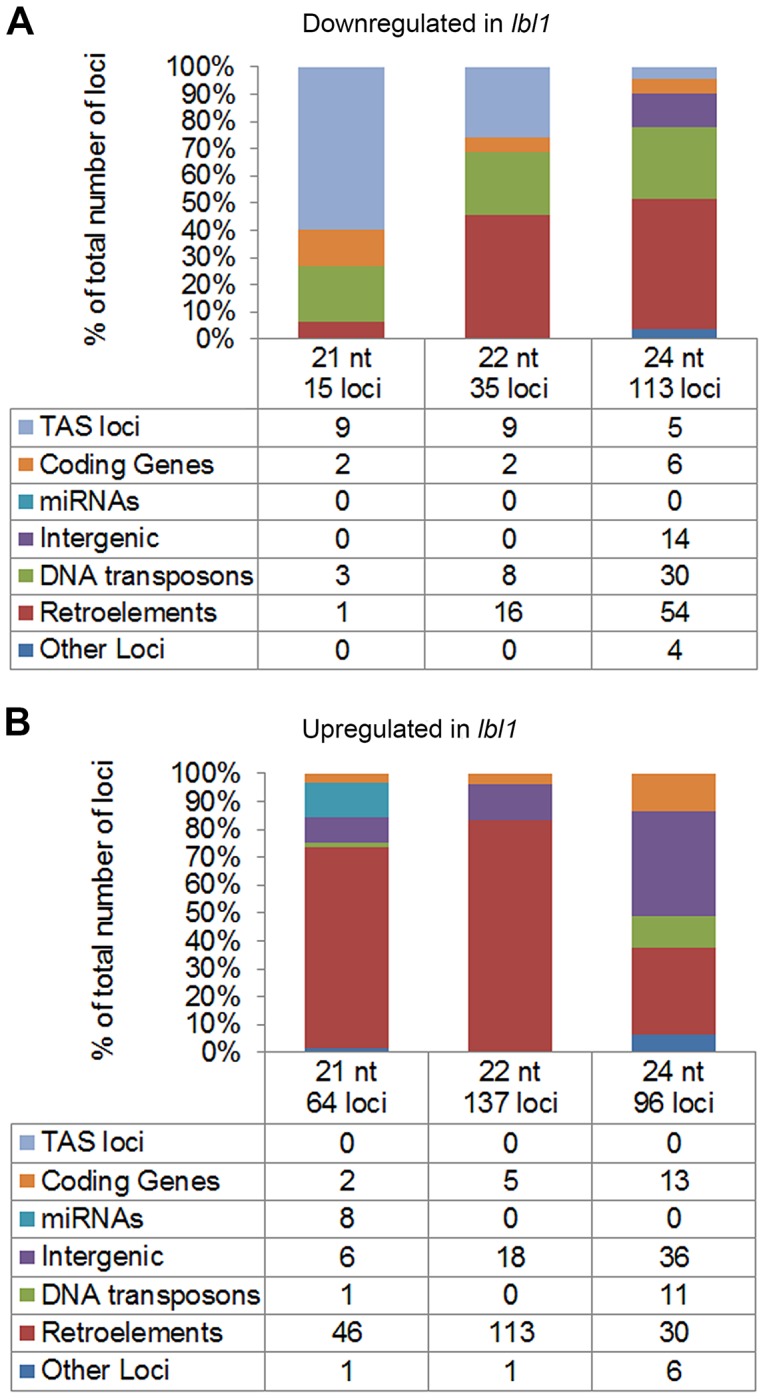
Maize leafbladeless1 (lbl1) encodes a key component in the trans-acting short-interfering RNA (ta-siRNA) biogenesis pathway. Correlated with a great diversity in ta-siRNAs and the targets they regulate, the phenotypes conditioned by mutants perturbing this small RNA pathway vary extensively across species. Mutations in lbl1 result in severe developmental defects, giving rise to plants with radial, abaxialized leaves. To investigate the basis for this phenotype, we compared the small RNA content between wild-type and lbl1 seedling apices. We show that LBL1 affects the accumulation of small RNAs in all major classes, and reveal unexpected crosstalk between ta-siRNA biogenesis and other small RNA pathways regulating transposons. Interestingly, in contrast to data from other plant species, we found no evidence for the existence of phased siRNAs generated via the one-hit model. Our analysis identified nine TAS loci, all belonging to the conserved TAS3 family. Information from RNA deep sequencing and PARE analyses identified the tasiR-ARFs as the major functional ta-siRNAs in the maize vegetative apex where they regulate expression of AUXIN RESPONSE FACTOR3 (ARF3) homologs. Plants expressing a tasiR-ARF insensitive arf3a transgene recapitulate the phenotype of lbl1, providing direct evidence that deregulation of ARF3 transcription factors underlies the developmental defects of maize ta-siRNA biogenesis mutants. The phenotypes of Arabidopsis and Medicago ta-siRNA mutants, while strikingly different, likewise result from misexpression of the tasiR-ARF target ARF3. Our data indicate that diversity in TAS pathways and their targets cannot fully account for the phenotypic differences conditioned by ta-siRNA biogenesis mutants across plant species. Instead, we propose that divergence in the gene networks downstream of the ARF3 transcription factors or the spatiotemporal pattern during leaf development in which these proteins act constitute key factors underlying the distinct contributions of the ta-siRNA pathway to development in maize, Arabidopsis, and possibly other plant species as well.
Conflict of interest statement
MCD and MCPT received salary support from Pioneer-DuPont. Pioneer-DuPont also generated the small RNA libraries. This does not alter our adherence to all PLOS policies on sharing data and materials.
Figures
Similar articles
-
Conserved LBL1-ta-siRNA and miR165/166-RLD1/2 modules regulate root development in maize.Development. 2021 Jan 5;148(1):dev190033. doi: 10.1242/dev.190033. Development. 2021. PMID: 33168582
-
Rice RNA-dependent RNA polymerase 6 acts in small RNA biogenesis and spikelet development.Plant J. 2012 Aug;71(3):378-89. doi: 10.1111/j.1365-313X.2012.05001.x. Epub 2012 Jun 5. Plant J. 2012. PMID: 22443269
-
Regulation of AUXIN RESPONSE FACTOR3 by TAS3 ta-siRNA affects developmental timing and patterning in Arabidopsis.Curr Biol. 2006 May 9;16(9):939-44. doi: 10.1016/j.cub.2006.03.065. Curr Biol. 2006. PMID: 16682356
-
Organ polarity in plants is specified through the opposing activity of two distinct small regulatory RNAs.Cold Spring Harb Symp Quant Biol. 2006;71:157-64. doi: 10.1101/sqb.2006.71.045. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381292 Review.
-
miRNAs in the biogenesis of trans-acting siRNAs in higher plants.Semin Cell Dev Biol. 2010 Oct;21(8):798-804. doi: 10.1016/j.semcdb.2010.03.008. Epub 2010 Mar 30. Semin Cell Dev Biol. 2010. PMID: 20359543 Review.
Cited by
-
Silencing of Mutator Elements in Maize Involves Distinct Populations of Small RNAs and Distinct Patterns of DNA Methylation.Genetics. 2020 Jun;215(2):379-391. doi: 10.1534/genetics.120.303033. Epub 2020 Mar 30. Genetics. 2020. PMID: 32229532 Free PMC article.
-
Extensive Families of miRNAs and PHAS Loci in Norway Spruce Demonstrate the Origins of Complex phasiRNA Networks in Seed Plants.Mol Biol Evol. 2015 Nov;32(11):2905-18. doi: 10.1093/molbev/msv164. Epub 2015 Aug 28. Mol Biol Evol. 2015. PMID: 26318183 Free PMC article.
-
Plant small RNAs: advancement in the understanding of biogenesis and role in plant development.Planta. 2018 Sep;248(3):545-558. doi: 10.1007/s00425-018-2927-5. Epub 2018 Jul 2. Planta. 2018. PMID: 29968061 Review.
-
Construction and Optimization of a Large Gene Coexpression Network in Maize Using RNA-Seq Data.Plant Physiol. 2017 Sep;175(1):568-583. doi: 10.1104/pp.17.00825. Epub 2017 Aug 2. Plant Physiol. 2017. PMID: 28768814 Free PMC article.
-
The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling.Noncoding RNA. 2024 Feb 7;10(1):13. doi: 10.3390/ncrna10010013. Noncoding RNA. 2024. PMID: 38392968 Free PMC article. Review.
References
-
- Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, et al. (2004) Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol Cell 16: 69–79. - PubMed
-
- Allen E, Xie Z, Gustafson A, Carrington J (2005) microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221. - PubMed
-
- Axtell M, Jan C, Rajagopalan R, Bartel D (2006) A two-hit trigger for siRNA biogenesis in plants. Cell 127: 565–577. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
