

Degradation of Cep68 and PCNT cleavage mediate Cep215 removal from the PCM to allow centriole separation, disengagement and licensing
- PMID: 25503564
- PMCID: PMC4415623
- DOI: 10.1038/ncb3076
Degradation of Cep68 and PCNT cleavage mediate Cep215 removal from the PCM to allow centriole separation, disengagement and licensing
Abstract
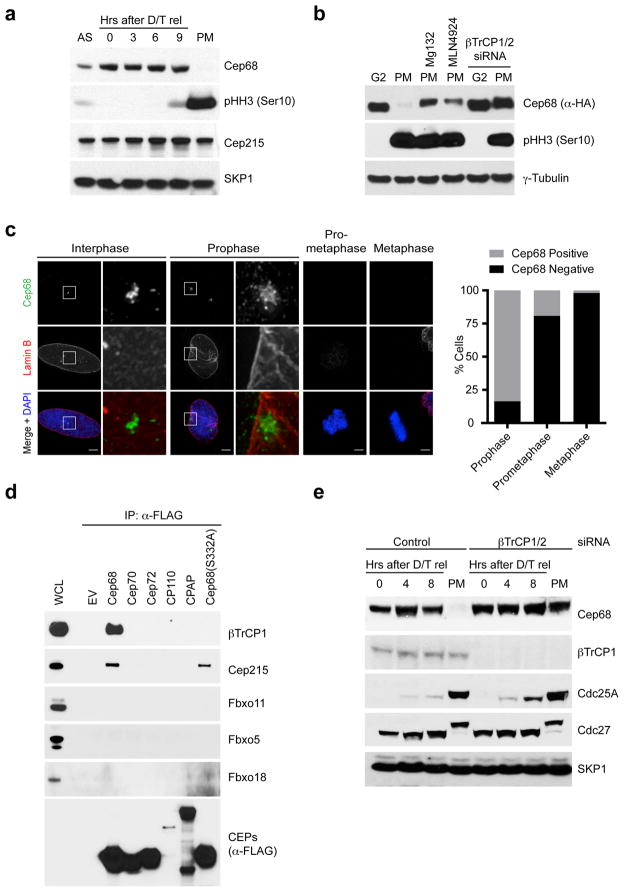
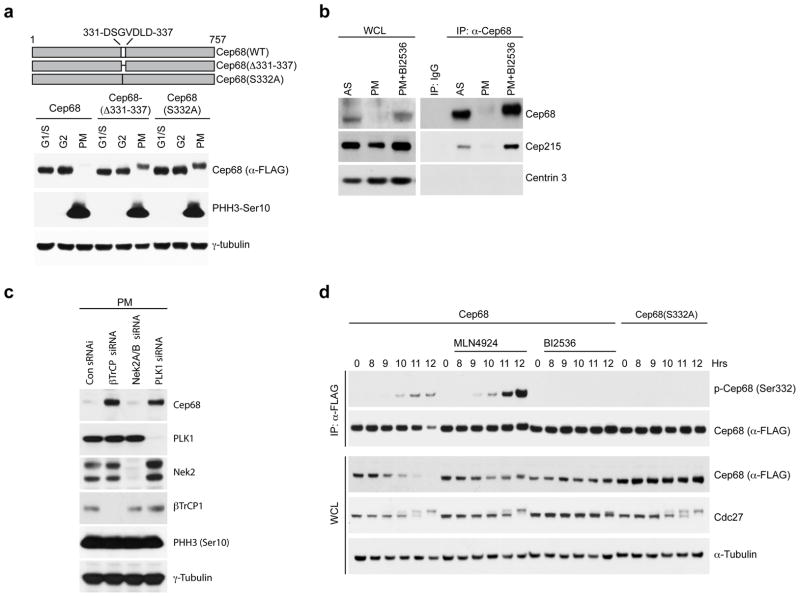
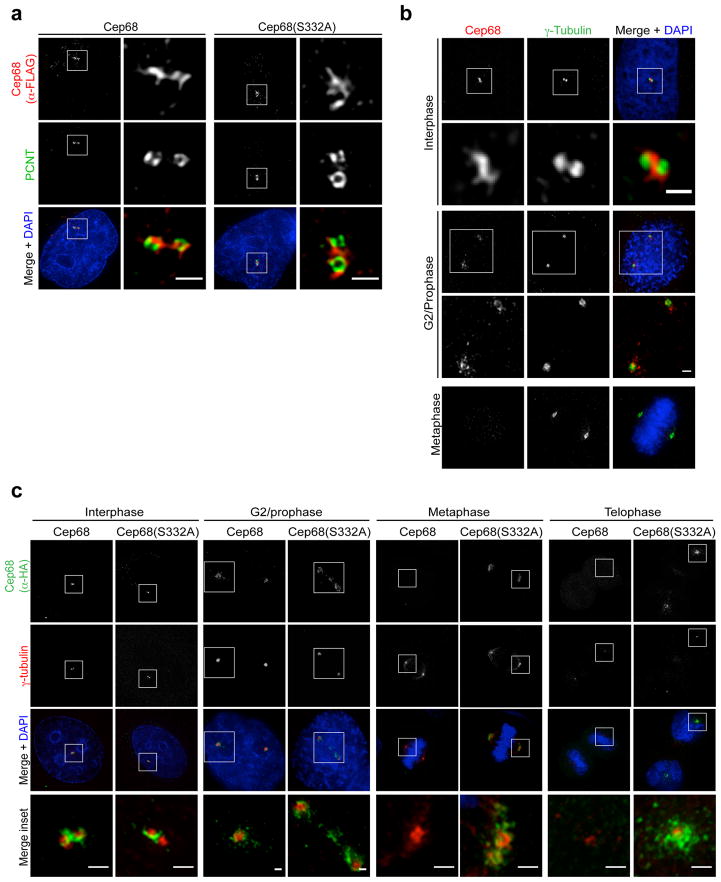
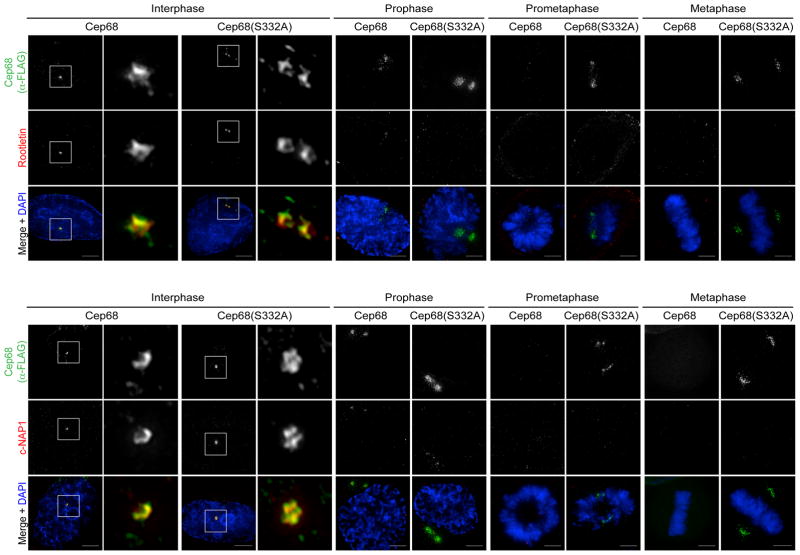
An intercentrosomal linker keeps a cell's two centrosomes joined together until it is dissolved at the onset of mitosis. A second connection keeps daughter centrioles engaged to their mothers until they lose their orthogonal arrangement at the end of mitosis. Centriole disengagement is required to license centrioles for duplication. We show that the intercentrosomal linker protein Cep68 is degraded in prometaphase through the SCF(βTrCP) (Skp1-Cul1-F-box protein) ubiquitin ligase complex. Cep68 degradation is initiated by PLK1 phosphorylation of Cep68 on Ser 332, allowing recognition by βTrCP. We also found that Cep68 forms a complex with Cep215 (also known as Cdk5Rap2) and PCNT (also known as pericentrin), two PCM (pericentriolar material) proteins involved in centriole engagement. Cep68 and PCNT bind to different pools of Cep215. We propose that Cep68 degradation allows Cep215 removal from the peripheral PCM preventing centriole separation following disengagement, whereas PCNT cleavage mediates Cep215 removal from the core of the PCM to inhibit centriole disengagement and duplication.
Figures
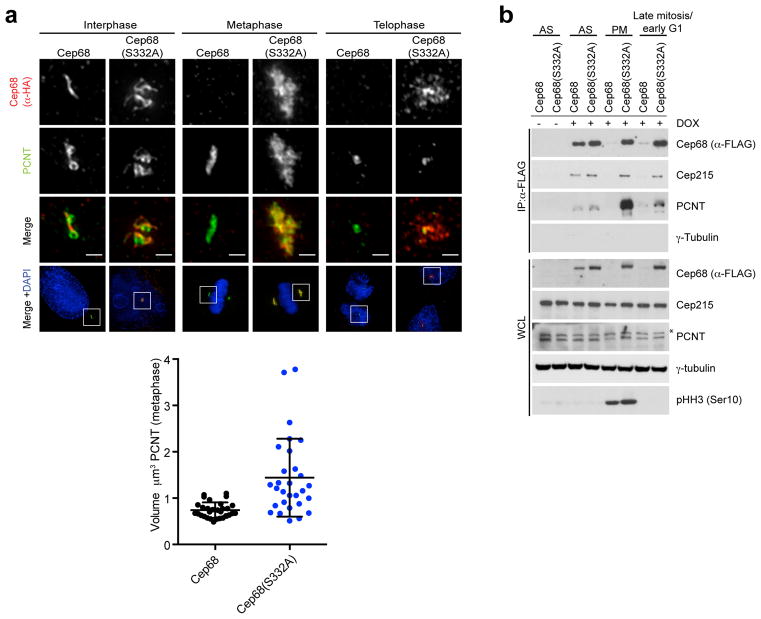
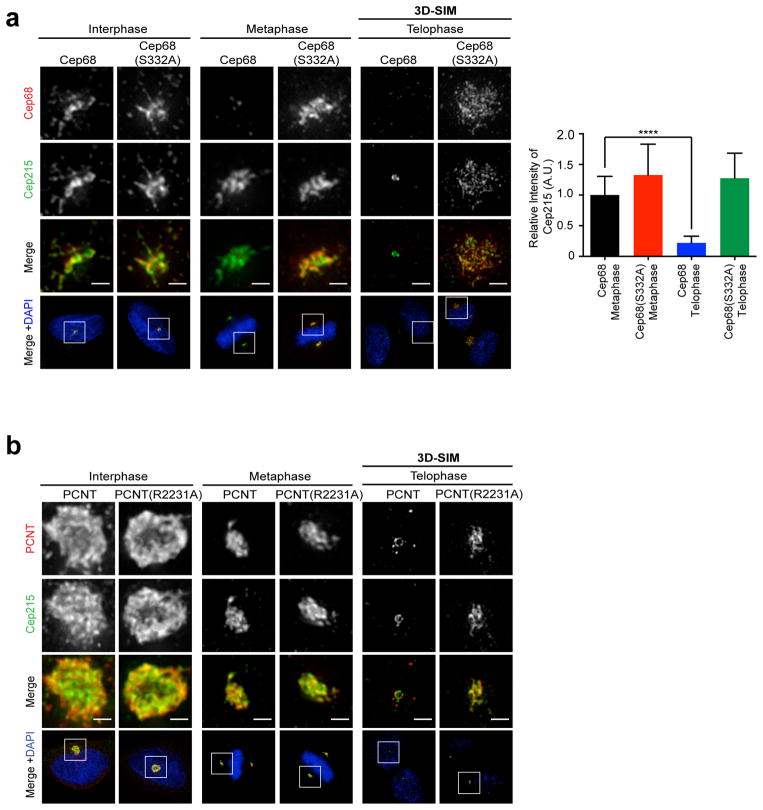
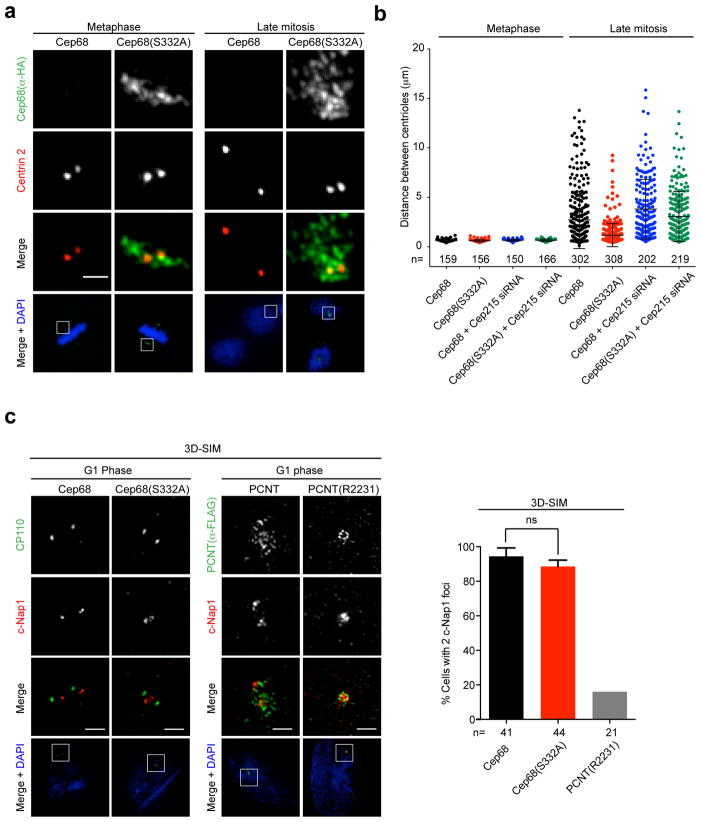
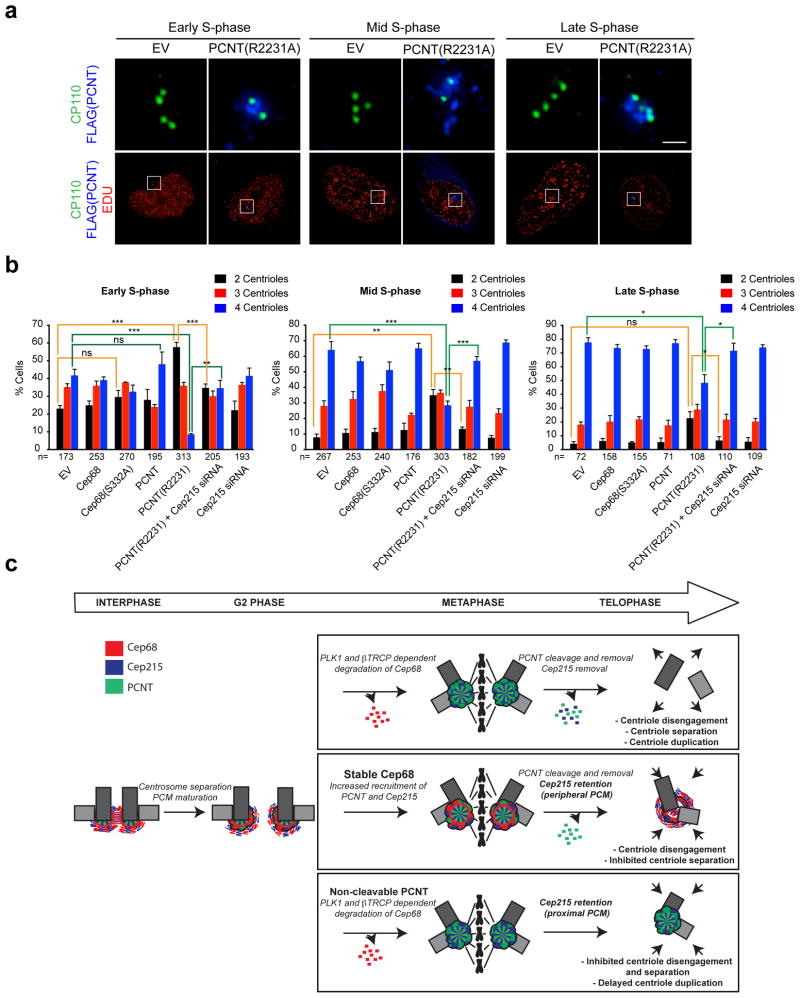
Comment in
-
Solving the centriole disengagement puzzle.Nat Cell Biol. 2015 Jan;17(1):3-5. doi: 10.1038/ncb3087. Nat Cell Biol. 2015. PMID: 25679029
References
-
- Bornens M, Paintrand M, Berges J, Marty MC, Karsenti E. Structural and chemical characterization of isolated centrosomes. Cell Motil Cytoskeleton. 1987;8:238–249. - PubMed
-
- Paintrand M, Moudjou M, Delacroix H, Bornens M. Centrosome organization and centriole architecture: their sensitivity to divalent cations. J Struct Biol. 1992;108:107–128. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
