

Systems biology. Accurate information transmission through dynamic biochemical signaling networks
- PMID: 25504722
- PMCID: PMC4813785
- DOI: 10.1126/science.1254933
Systems biology. Accurate information transmission through dynamic biochemical signaling networks
Abstract
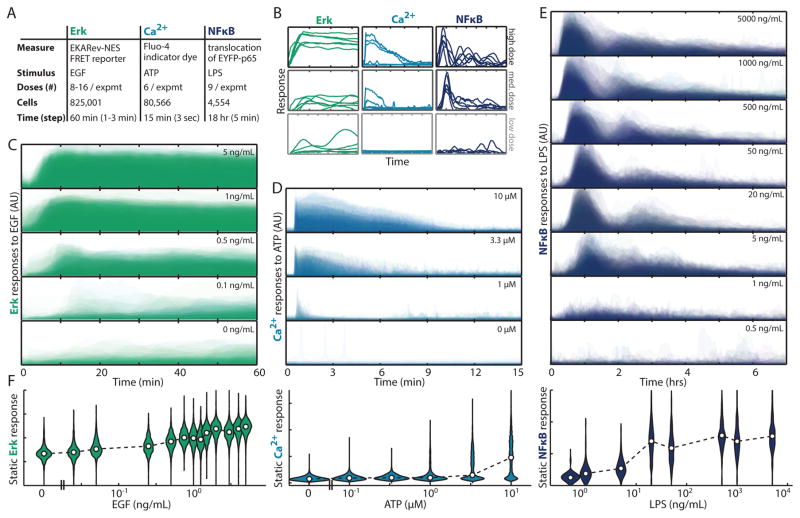
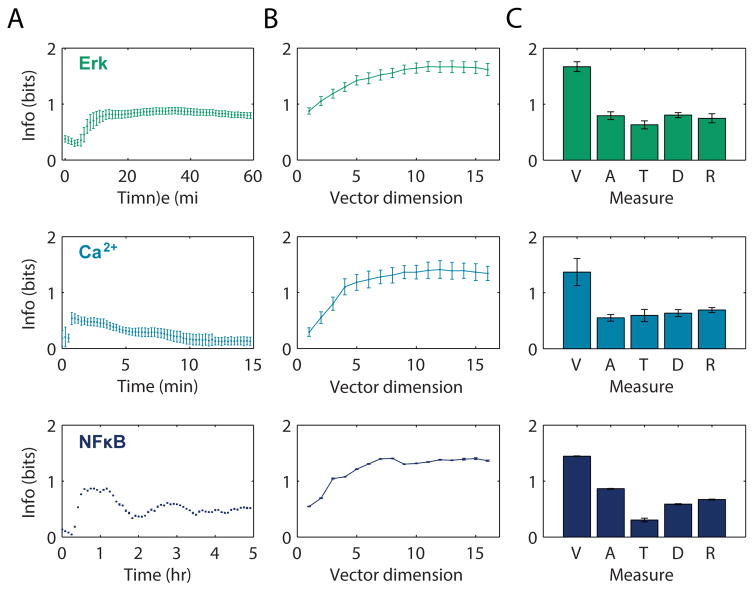
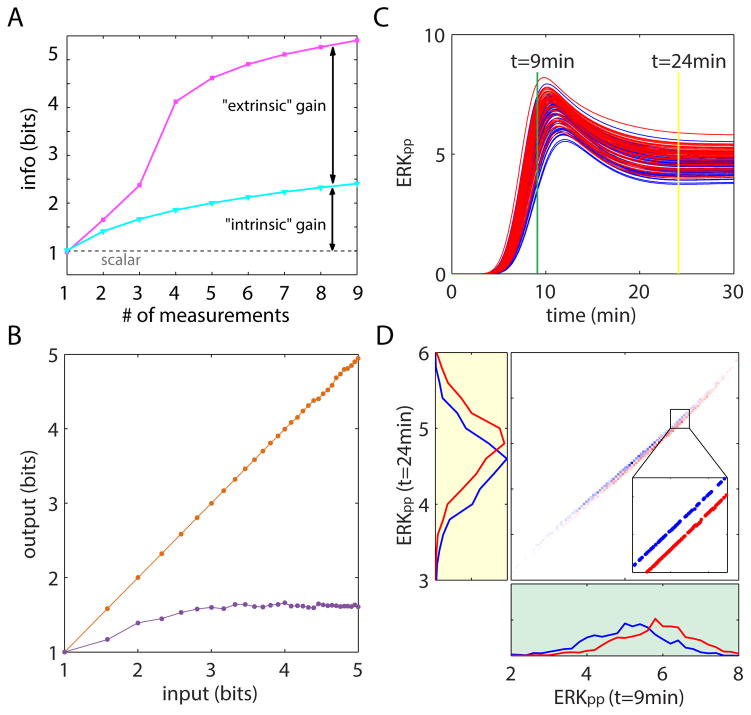
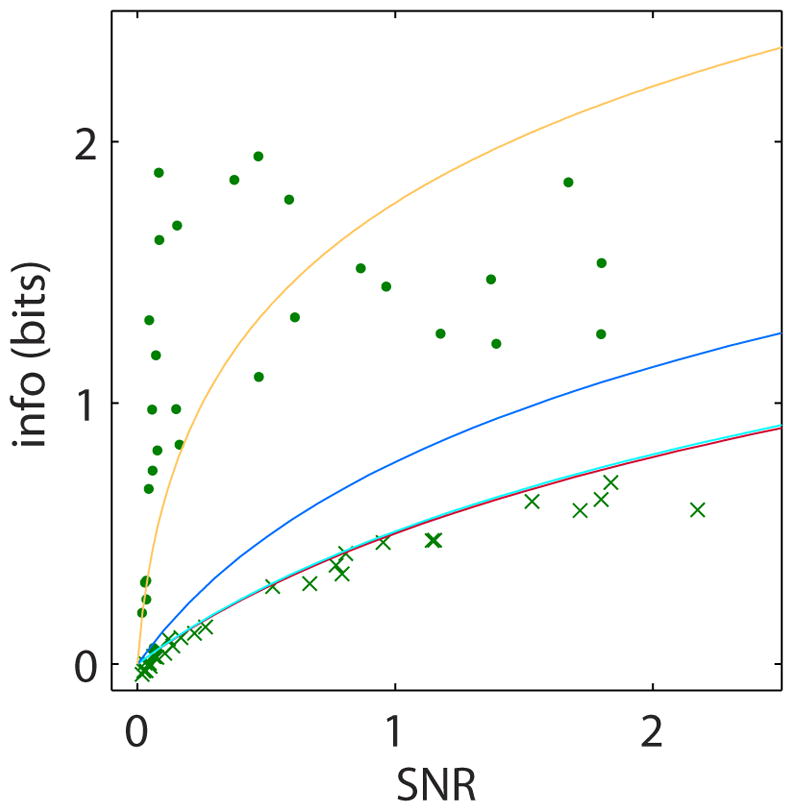
Stochasticity inherent to biochemical reactions (intrinsic noise) and variability in cellular states (extrinsic noise) degrade information transmitted through signaling networks. We analyzed the ability of temporal signal modulation--that is, dynamics--to reduce noise-induced information loss. In the extracellular signal-regulated kinase (ERK), calcium (Ca(2+)), and nuclear factor kappa-B (NF-κB) pathways, response dynamics resulted in significantly greater information transmission capacities compared to nondynamic responses. Theoretical analysis demonstrated that signaling dynamics has a key role in overcoming extrinsic noise. Experimental measurements of information transmission in the ERK network under varying signal-to-noise levels confirmed our predictions and showed that signaling dynamics mitigate, and can potentially eliminate, extrinsic noise-induced information loss. By curbing the information-degrading effects of cell-to-cell variability, dynamic responses substantially increase the accuracy of biochemical signaling networks.
Copyright © 2014, American Association for the Advancement of Science.
Figures
References
-
- Waltermann C, Klipp E. Information theory based approaches to cellular signaling. Biochim Biophys Acta. 2011;1810:924–32. - PubMed
-
- Tostevin F, ten Wolde PR. Mutual Information between Input and Output Trajectories of Biochemical Networks. Phys Rev Lett. 2009;102:218101. - PubMed
-
- de Ronde WH, Tostevin F, ten Wolde PR. Effect of feedback on the fidelity of information transmission of time-varying signals. Phys Rev E. 2010;82:031914. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
