

Collective sensing and collective responses in quorum-sensing bacteria
- PMID: 25505130
- PMCID: PMC4305403
- DOI: 10.1098/rsif.2014.0882
Collective sensing and collective responses in quorum-sensing bacteria
Abstract
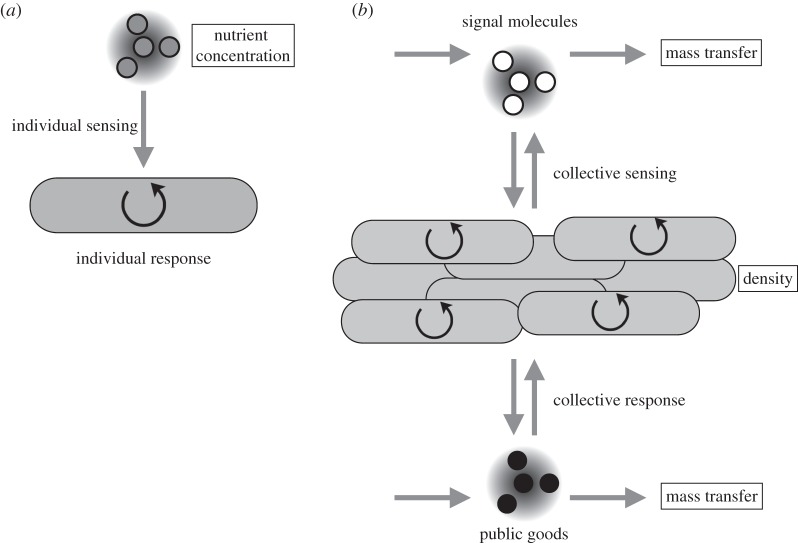
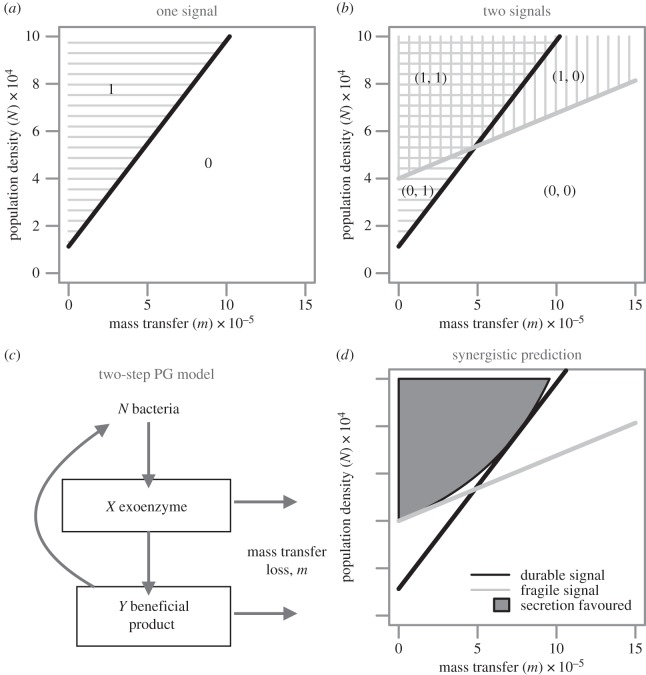
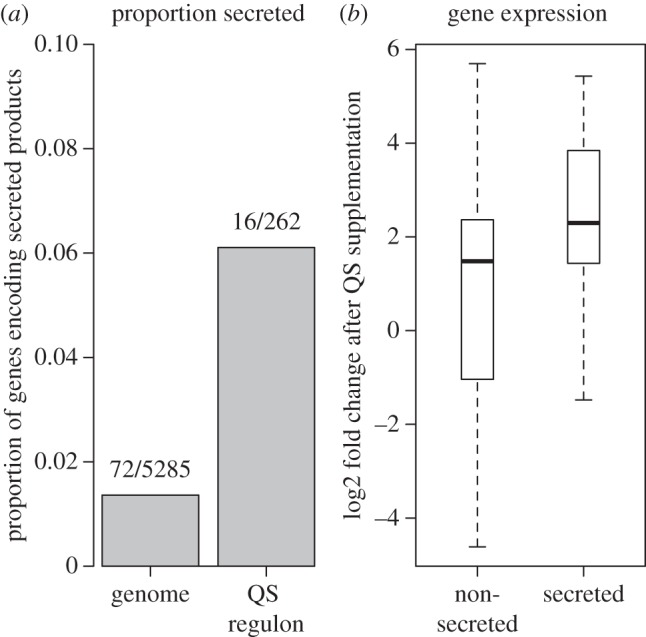
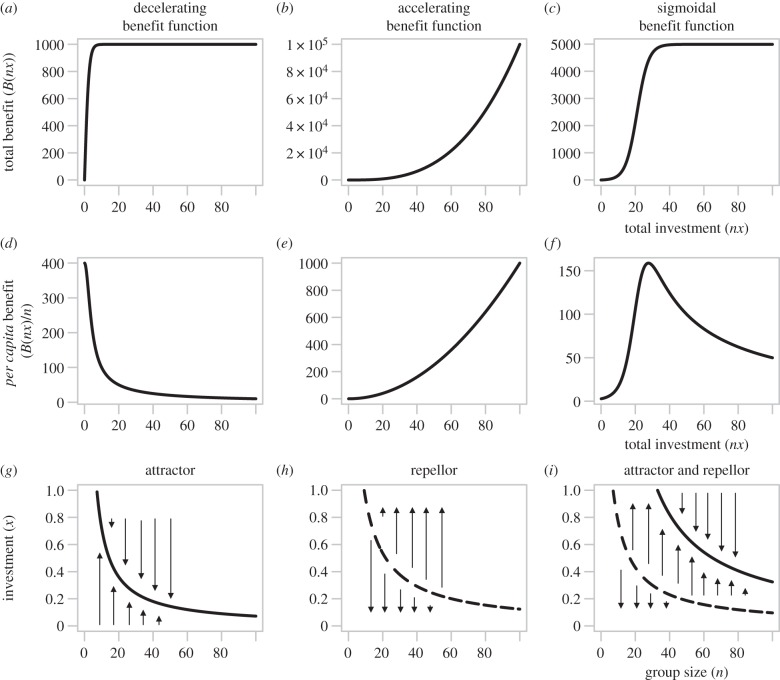
Bacteria often face fluctuating environments, and in response many species have evolved complex decision-making mechanisms to match their behaviour to the prevailing conditions. Some environmental cues provide direct and reliable information (such as nutrient concentrations) and can be responded to individually. Other environmental parameters are harder to infer and require a collective mechanism of sensing. In addition, some environmental challenges are best faced by a group of cells rather than an individual. In this review, we discuss how bacteria sense and overcome environmental challenges as a group using collective mechanisms of sensing, known as 'quorum sensing' (QS). QS is characterized by the release and detection of small molecules, potentially allowing individuals to infer environmental parameters such as density and mass transfer. While a great deal of the molecular mechanisms of QS have been described, there is still controversy over its functional role. We discuss what QS senses and how, what it controls and why, and how social dilemmas shape its evolution. Finally, there is a growing focus on the use of QS inhibitors as antibacterial chemotherapy. We discuss the claim that such a strategy could overcome the evolution of resistance. By linking existing theoretical approaches to data, we hope this review will spur greater collaboration between experimental and theoretical researchers.
Keywords: collective behaviour; quorum sensing; social evolution; systems biology.
Figures
References
-
- Pardee AB, Jacob F, Monod J. 1959. The genetic control and cytoplasmic expression of ‘inducibility’ in the synthesis of β-galactosidase by E. coli. J. Mol. Biol. 1, 165–178. ( 10.1016/S0022-2836(59)80045-0) - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources