

Heart mitochondrial TTP synthesis and the compartmentalization of TMP
- PMID: 25505243
- PMCID: PMC4303658
- DOI: 10.1074/jbc.M114.624213
Heart mitochondrial TTP synthesis and the compartmentalization of TMP
Abstract
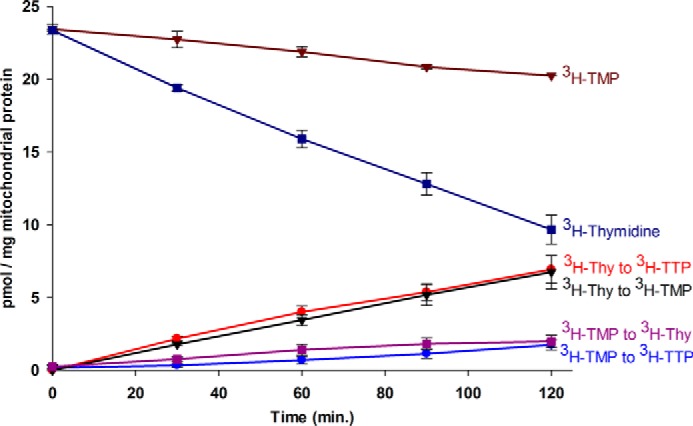
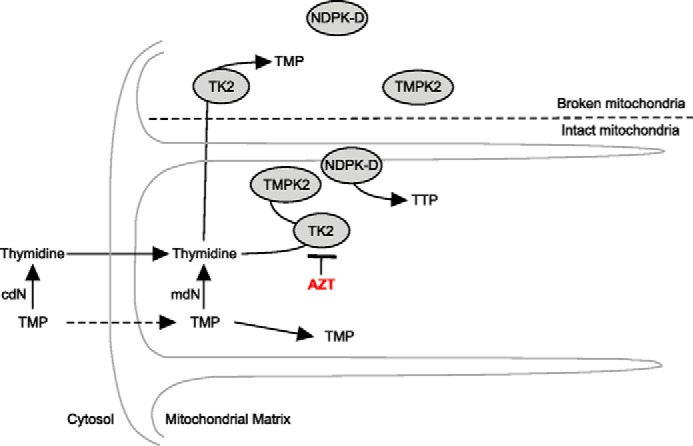
The primary pathway of TTP synthesis in the heart requires thymidine salvage by mitochondrial thymidine kinase 2 (TK2). However, the compartmentalization of this pathway and the transport of thymidine nucleotides are not well understood. We investigated the metabolism of [(3)H]thymidine or [(3)H]TMP as precursors of [(3)H]TTP in isolated intact or broken mitochondria from the rat heart. The results demonstrated that [(3)H]thymidine was readily metabolized by the mitochondrial salvage enzymes to TTP in intact mitochondria. The equivalent addition of [(3)H]TMP produced far less [(3)H]TTP than the amount observed with [(3)H]thymidine as the precursor. Using zidovudine to inhibit TK2, the synthesis of [(3)H]TTP from [(3)H]TMP was effectively blocked, demonstrating that synthesis of [(3)H]TTP from [(3)H]TMP arose solely from the dephosphorysynthase pathway that includes deoxyuridine triphosphatelation of [(3)H]TMP to [(3)H]thymidine. To determine the role of the membrane in TMP metabolism, mitochondrial membranes were disrupted by freezing and thawing. In broken mitochondria, [(3)H]thymidine was readily converted to [(3)H]TMP, but further phosphorylation was prevented even though the energy charge was well maintained by addition of oligomycin A, phosphocreatine, and creatine phosphokinase. The failure to synthesize TTP in broken mitochondria was not related to a loss of membrane potential or inhibition of the electron transport chain, as confirmed by addition of carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone and potassium cyanide, respectively, in intact mitochondria. In summary, these data, taken together, suggest that the thymidine salvage pathway is compartmentalized so that TMP kinase prefers TMP synthesized by TK2 over medium TMP and that this is disrupted in broken mitochondria.
Keywords: Mitochondria; Mitochondrial Metabolism; Nucleoside/Nucleotide Biosynthesis; Nucleoside/Nucleotide Metabolism; Thymidine Kinase 2.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
