

Independent premotor encoding of the sequence and structure of birdsong in avian cortex
- PMID: 25505334
- PMCID: PMC6608506
- DOI: 10.1523/JNEUROSCI.1940-14.2014
Independent premotor encoding of the sequence and structure of birdsong in avian cortex
Abstract
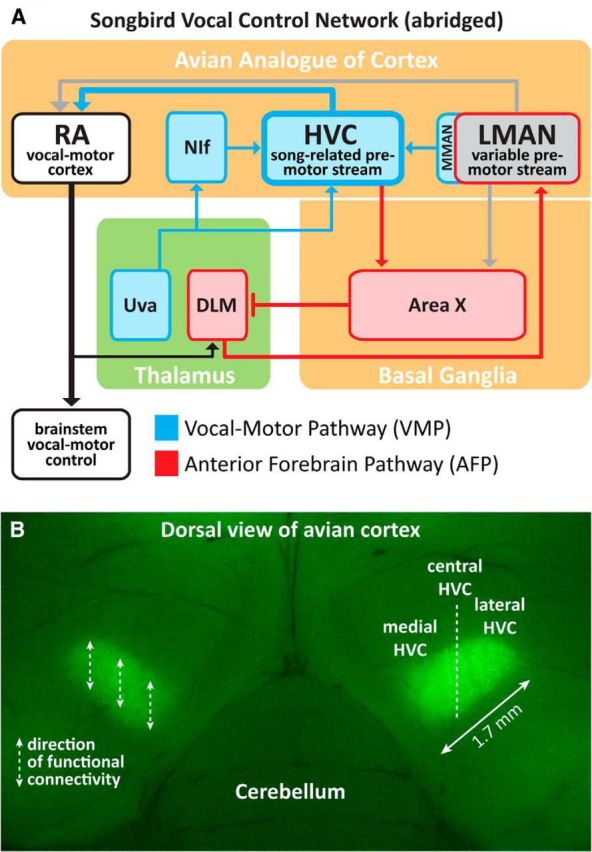
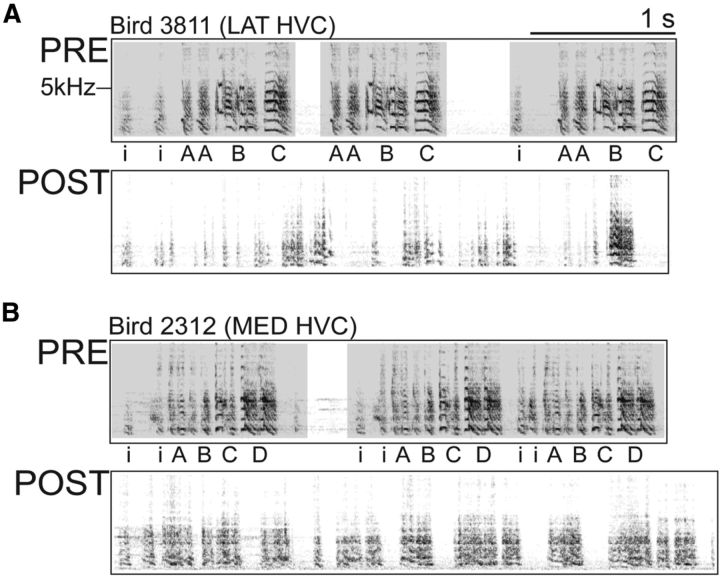
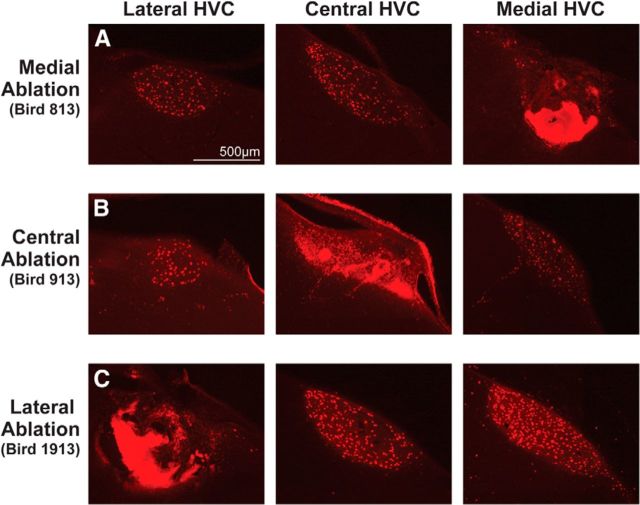
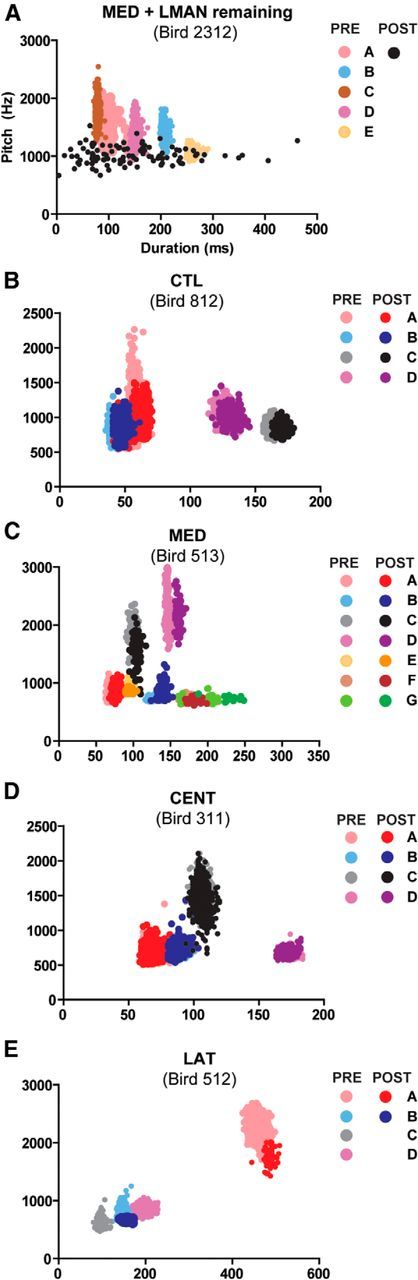
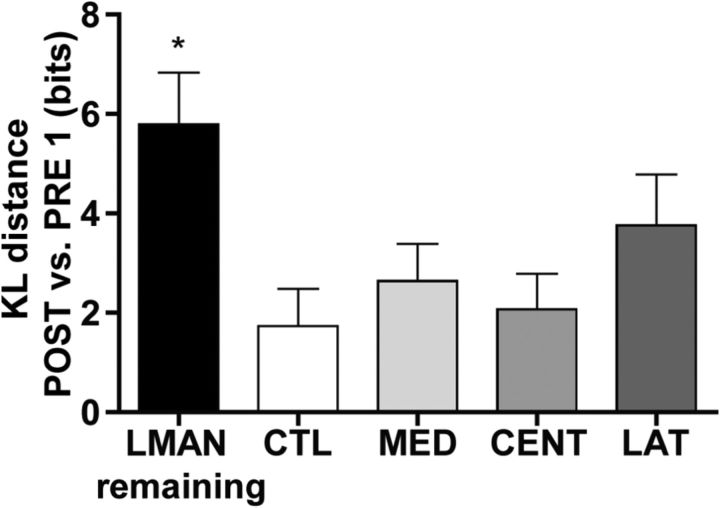
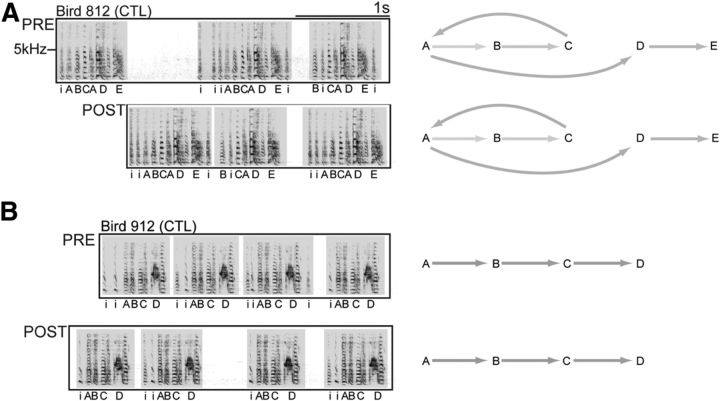
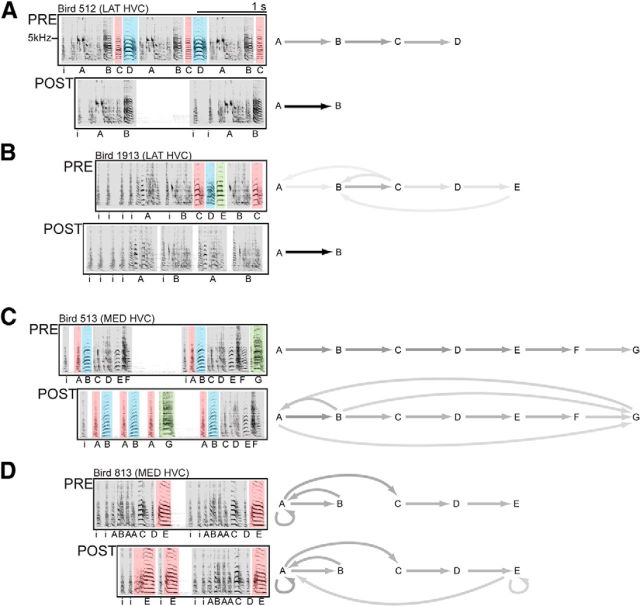
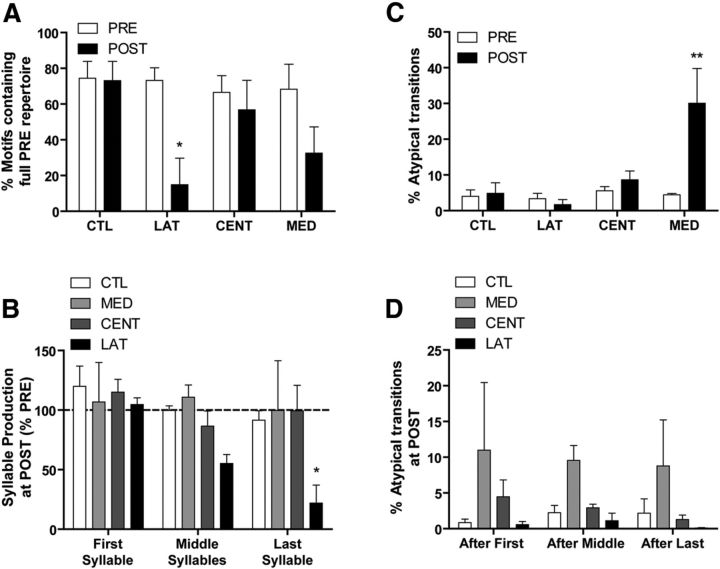
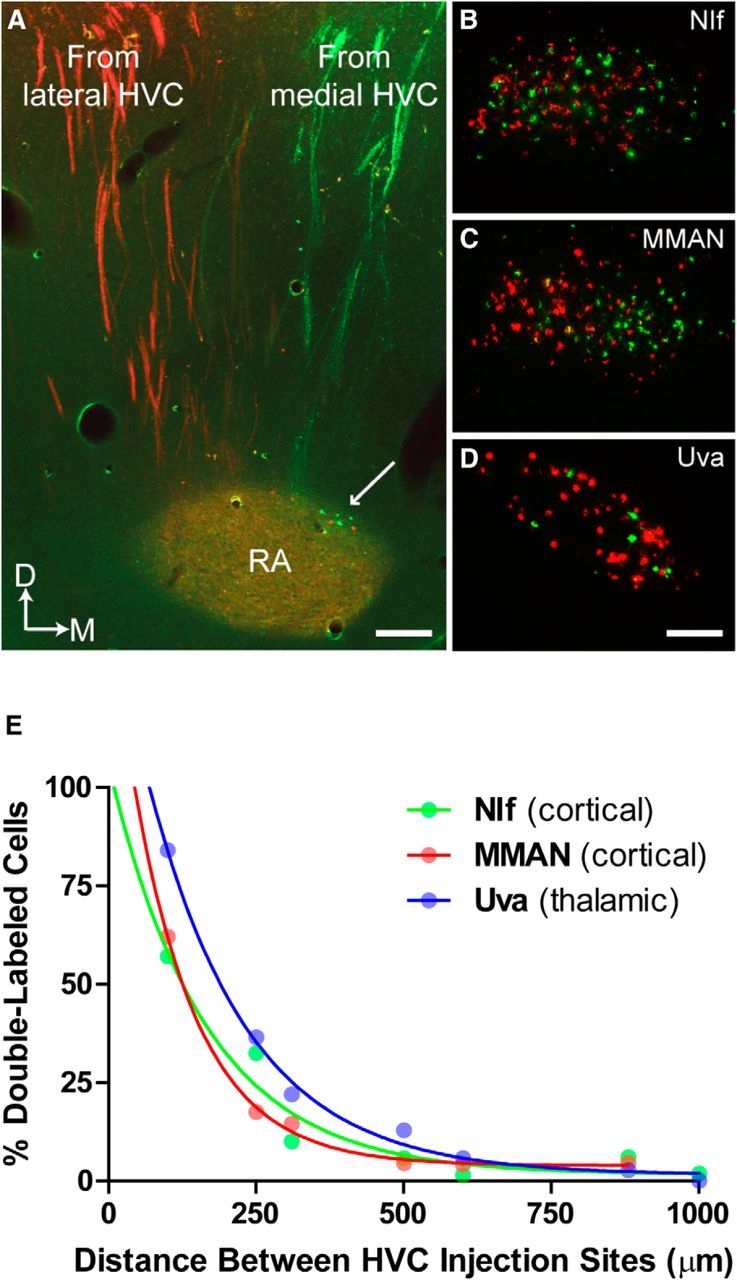
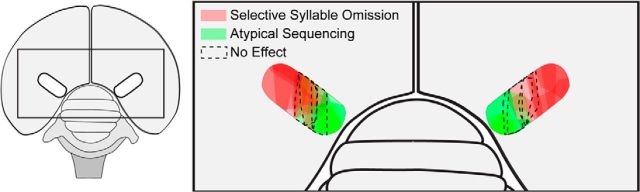
How the brain coordinates rapid sequences of learned behavior, such as human speech, remains a fundamental problem in neuroscience. Birdsong is a model of such behavior, which is learned and controlled by a neural circuit that spans avian cortex, basal ganglia, and thalamus. The songs of adult male zebra finches (Taeniopygia guttata), produced as rapid sequences of vocal gestures (syllables), are encoded by the cortical premotor region HVC (proper name). While the motor encoding of song within HVC has traditionally been viewed as unitary and distributed, we used an ablation technique to ask whether the sequence and structure of song are processed independently within HVC. Results revealed a functional topography across the medial-lateral axis of HVC. Bilateral ablation of medial HVC induced a positive disruption of song (increase in atypical syllable sequences), whereas bilateral ablation of lateral HVC induced a negative disruption (omission of individual syllables). Bilateral ablation of central HVC either had no effect on song or induced syllable omission, similar to lateral HVC ablation. We then investigated HVC connectivity and found parallel afferent and efferent pathways that transit medial and lateral HVC and converge at vocal motor cortex. In light of recent evidence that syntactic and lexical components of human speech are processed independently by neighboring regions of cortex (Menenti et al., 2012), our demonstration of anatomically distinct pathways that differentially process the sequence and structure of birdsong in parallel suggests that the vertebrate brain relies on a common approach to encode rapid sequences of vocal gestures.
Keywords: ablation; motor encoding; parallel processing; serial-order behavior; tract-tracing; zebra finch.
Copyright © 2014 the authors 0270-6474/14/3416821-14$15.00/0.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources