

Transposable element islands facilitate adaptation to novel environments in an invasive species
- PMID: 25510865
- PMCID: PMC4284661
- DOI: 10.1038/ncomms6495
Transposable element islands facilitate adaptation to novel environments in an invasive species
Abstract
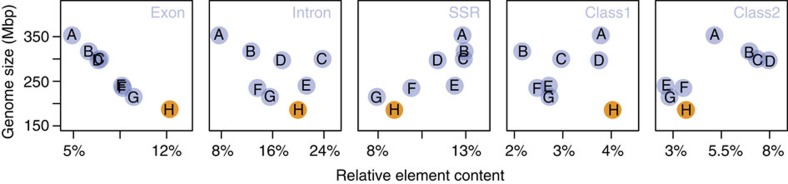
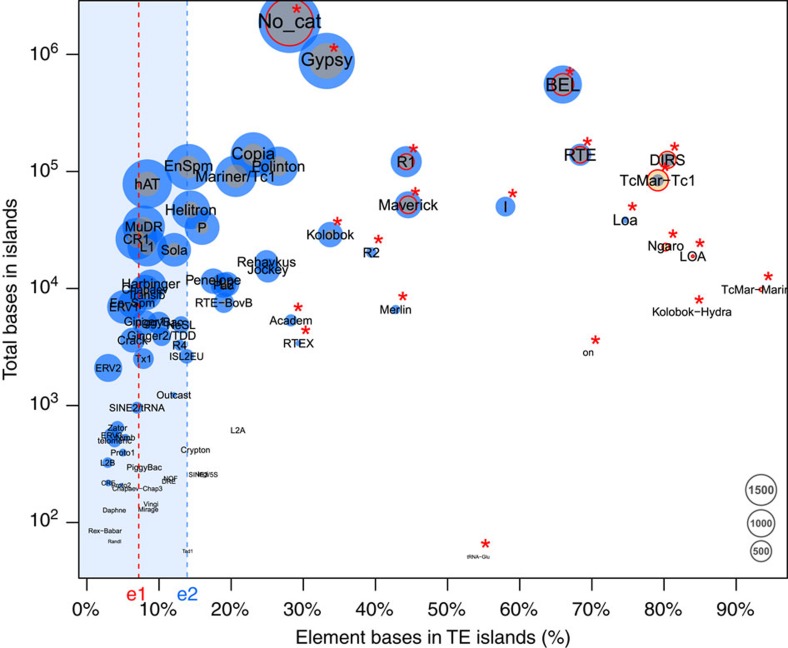
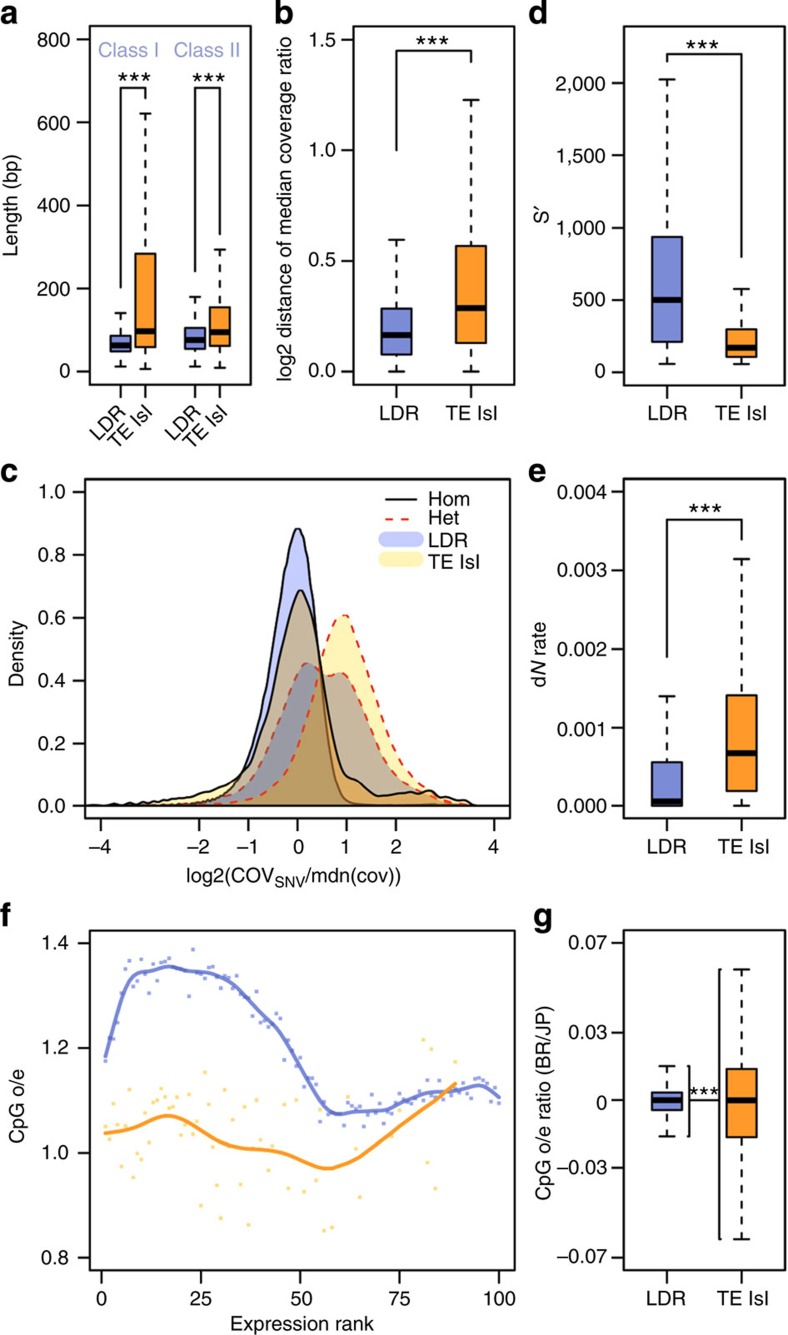
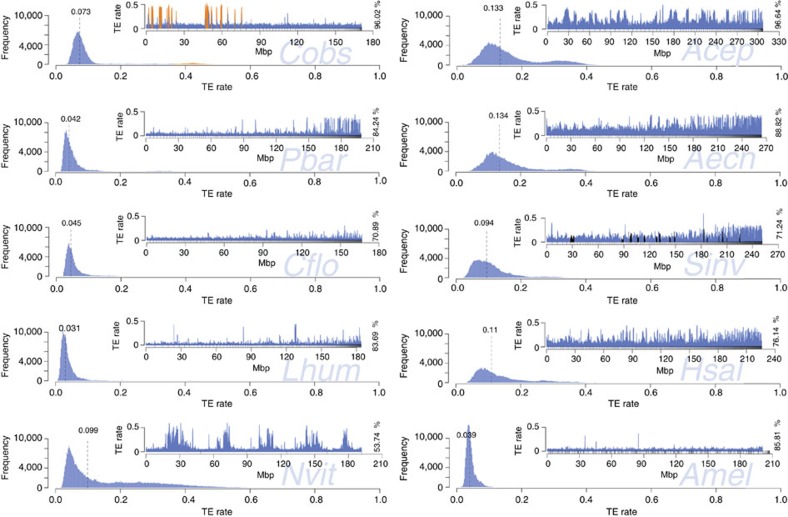
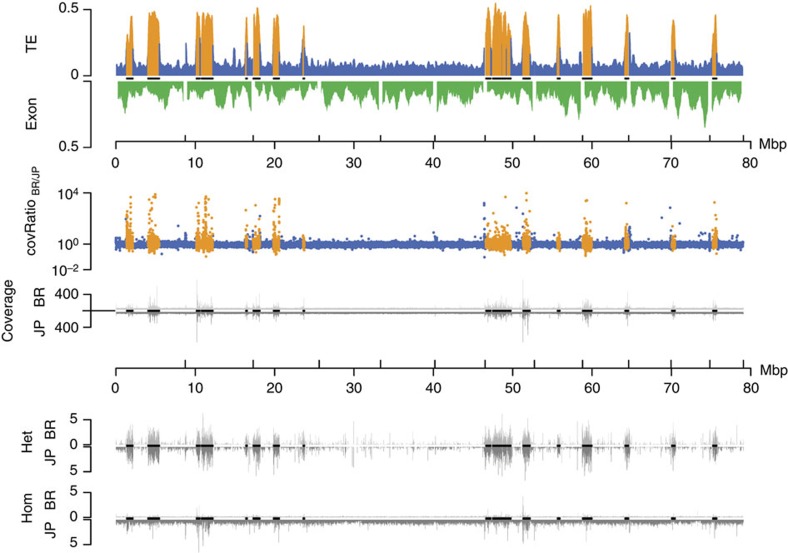
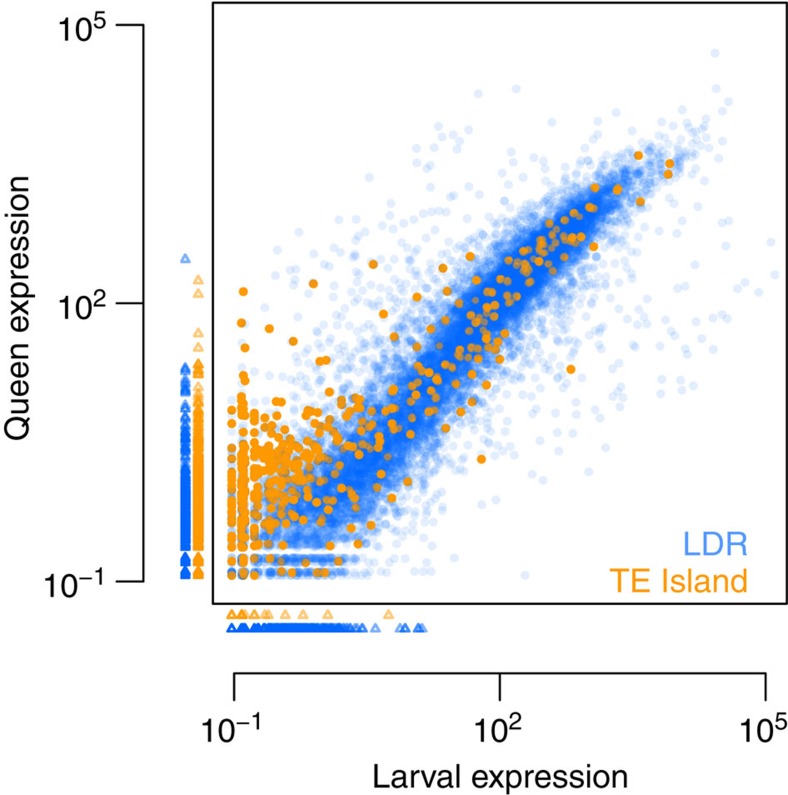
Adaptation requires genetic variation, but founder populations are generally genetically depleted. Here we sequence two populations of an inbred ant that diverge in phenotype to determine how variability is generated. Cardiocondyla obscurior has the smallest of the sequenced ant genomes and its structure suggests a fundamental role of transposable elements (TEs) in adaptive evolution. Accumulations of TEs (TE islands) comprising 7.18% of the genome evolve faster than other regions with regard to single-nucleotide variants, gene/exon duplications and deletions and gene homology. A non-random distribution of gene families, larvae/adult specific gene expression and signs of differential methylation in TE islands indicate intragenomic differences in regulation, evolutionary rates and coalescent effective population size. Our study reveals a tripartite interplay between TEs, life history and adaptation in an invasive species.
Figures

References
-
- Charlesworth D. & Charlesworth B. Inbreeding depression and its evolutionary consequences. Annu. Rev. Ecol. Syst. 18, 237–268 (1987).
-
- Lynch M. The Origins of Genome Architecture Sinauer Associates Inc (2007).
-
- Charlesworth D. & Wright S. I. Breeding systems and genome evolution. Curr. Opin. Genet. Dev. 11, 685–690 (2001). - PubMed
-
- Romiguier J. et al. Population genomics of eusocial insects: the costs of a vertebrate-like effective population size. J. Evol. Biol. 27, 593–603 (2014). - PubMed
-
- McDonald J. H. & Kreitman M. Adaptive protein evolution at the Adh locus in Drosophila. Nature 351, 652–654 (1991). - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
