

Biochemical principles underlying the stable maintenance of LTP by the CaMKII/NMDAR complex
- PMID: 25511992
- PMCID: PMC4466085
- DOI: 10.1016/j.brainres.2014.12.010
Biochemical principles underlying the stable maintenance of LTP by the CaMKII/NMDAR complex
Abstract
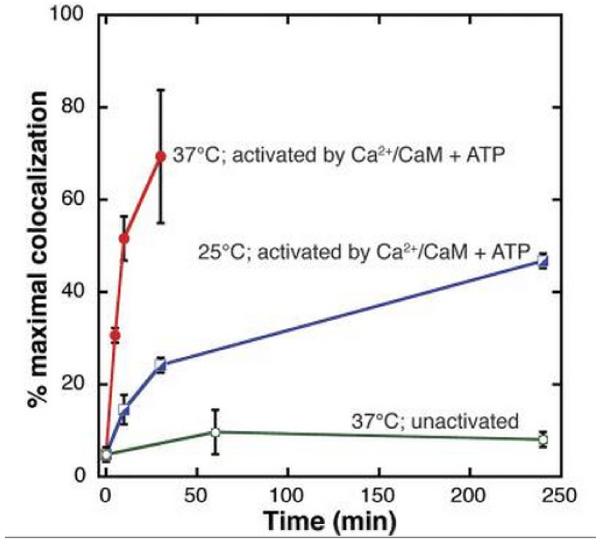
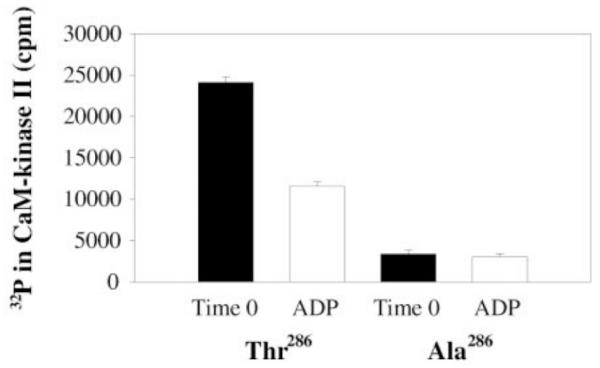
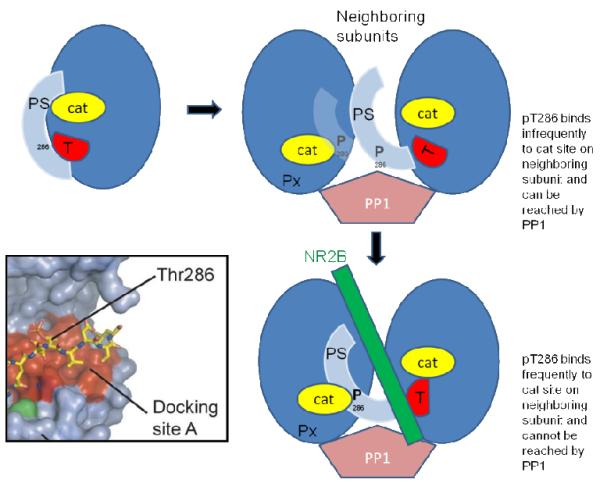
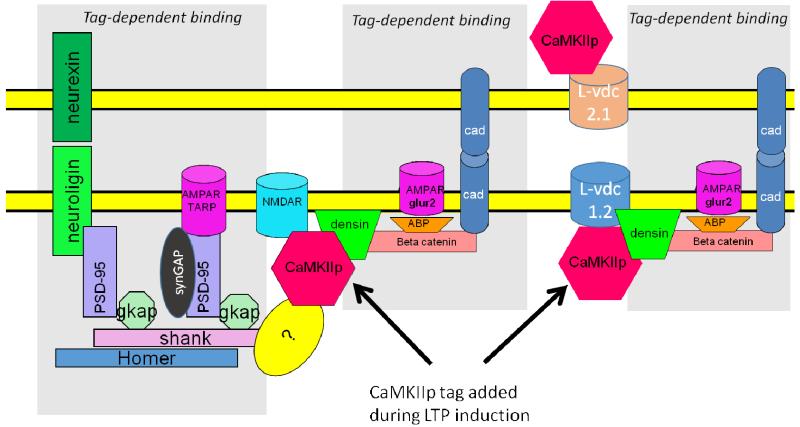
Memory involves the storage of information at synapses by an LTP-like process. This information storage is synapse specific and can endure for years despite the turnover of all synaptic proteins. There must, therefore, be special principles that underlie the stability of LTP. Recent experimental results suggest that LTP is maintained by the complex of CaMKII with the NMDAR. Here we consider the specifics of the CaMKII/NMDAR molecular switch, with the goal of understanding the biochemical principles that underlie stable information storage by synapses. Consideration of a variety of experimental results suggests that multiple principles are involved. One switch requirement is to prevent spontaneous transitions from the off to the on state. The highly cooperative nature of CaMKII autophosphorylation by Ca(2+) (Hill coefficient of 8) and the fact that formation of the CaMKII/NMDAR complex requires release of CaMKII from actin are mechanisms that stabilize the off state. The stability of the on state depends critically on intersubunit autophosphorylation, a process that restores any loss of pT286 due to phosphatase activity. Intersubunit autophosphorylation is also important in explaining why on state stability is not compromised by protein turnover. Recent evidence suggests that turnover occurs by subunit exchange. Thus, stability could be achieved if a newly inserted unphosphorylated subunit was autophosphorylated by a neighboring subunit. Based on other recent work, we posit a novel mechanism that enhances the stability of the on state by protection of pT286 from phosphatases. We posit that the binding of the NMNDAR to CaMKII forces pT286 into the catalytic site of a neighboring subunit, thereby protecting pT286 from phosphatases. A final principle concerns the role of structural changes. The binding of CaMKII to the NMDAR may act as a tag to organize the binding of further proteins that produce the synapse enlargement that underlies late LTP. We argue that these structural changes not only enhance transmission, but also enhance the stability of the CaMKII/NMDAR complex. Together, these principles provide a mechanistic framework for understanding how individual synapses produce stable information storage. This article is part of a Special Issue entitled SI: Brain and Memory.
Keywords: Kinase; LTP; Memory; Phosphatase.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
