

Identification of S-nitroso-CoA reductases that regulate protein S-nitrosylation
- PMID: 25512491
- PMCID: PMC4284529
- DOI: 10.1073/pnas.1417816112
Identification of S-nitroso-CoA reductases that regulate protein S-nitrosylation
Abstract
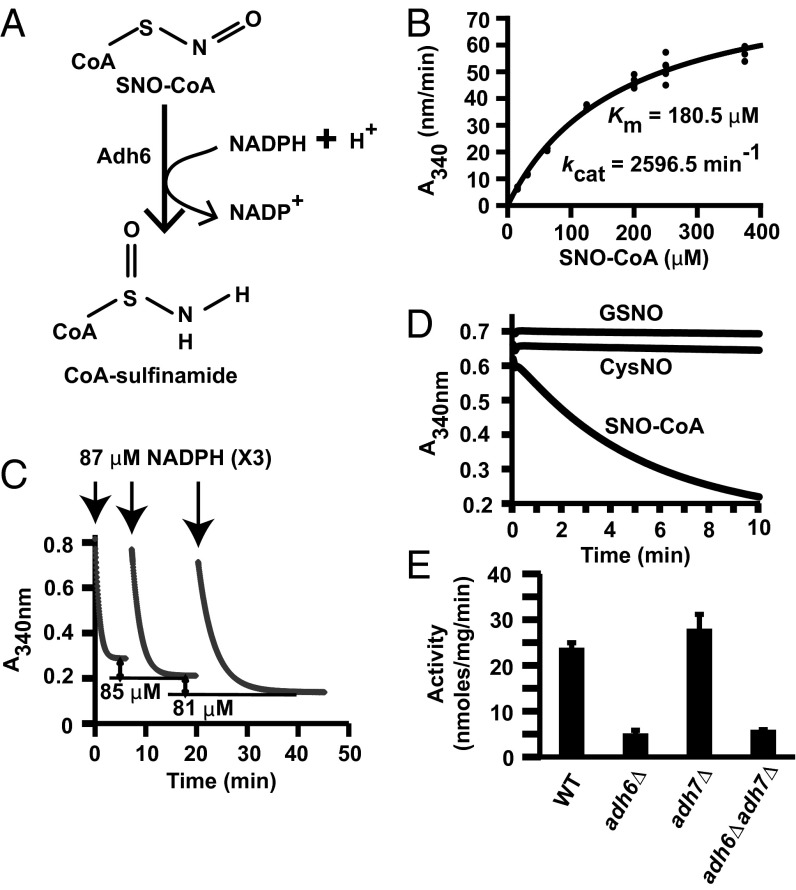
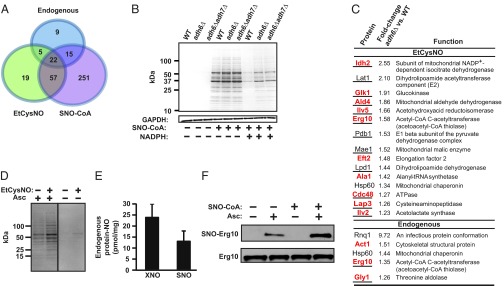
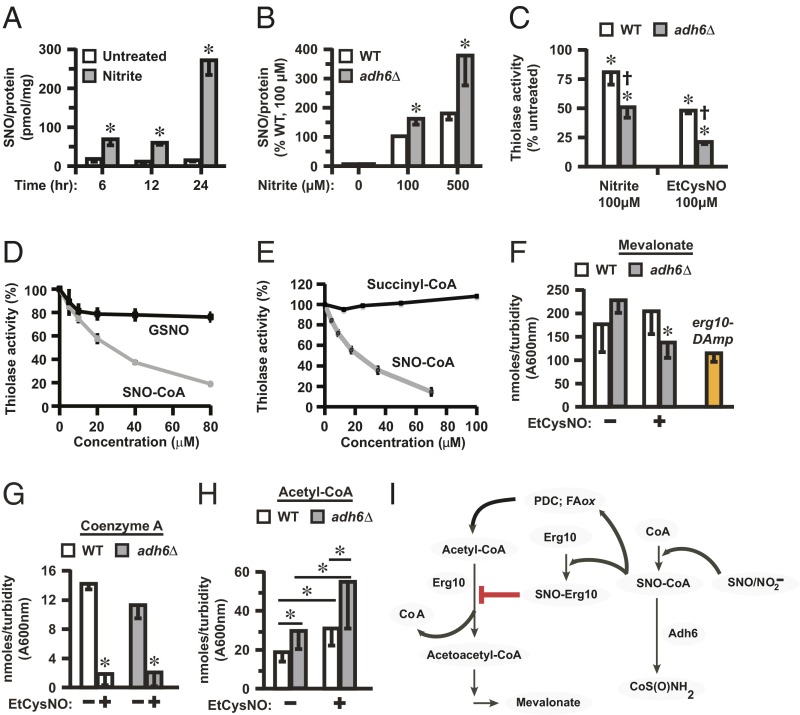
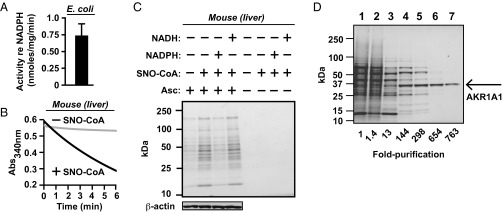
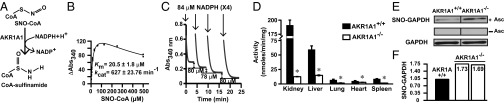
Coenzyme A (CoA) mediates thiol-based acyl-group transfer (acetylation and palmitoylation). However, a role for CoA in the thiol-based transfer of NO groups (S-nitrosylation) has not been considered. Here we describe protein S-nitrosylation in yeast (heretofore unknown) that is mediated by S-nitroso-CoA (SNO-CoA). We identify a specific SNO-CoA reductase encoded by the alcohol dehydrogenase 6 (ADH6) gene and show that deletion of ADH6 increases cellular S-nitrosylation and alters CoA metabolism. Further, we report that Adh6, acting as a selective SNO-CoA reductase, protects acetoacetyl-CoA thiolase from inhibitory S-nitrosylation and thereby affects sterol biosynthesis. Thus, Adh6-regulated, SNO-CoA-mediated protein S-nitrosylation provides a regulatory mechanism paralleling protein acetylation. We also find that SNO-CoA reductases are present from bacteria to mammals, and we identify aldo-keto reductase 1A1 as the mammalian functional analog of Adh6. Our studies reveal a novel functional class of enzymes that regulate protein S-nitrosylation from yeast to mammals and suggest that SNO-CoA-mediated S-nitrosylation may subserve metabolic regulation.
Keywords: AKR1A1; Adh6; S-nitrosylation; denitrosylase; denitrosylation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Benhar M, Forrester MT, Stamler JS. Protein denitrosylation: Enzymatic mechanisms and cellular functions. Nat Rev Mol Cell Biol. 2009;10(10):721–732. - PubMed
-
- Liu L, et al. A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature. 2001;410(6827):490–494. - PubMed
-
- Liu L, et al. Essential roles of S-nitrosothiols in vascular homeostasis and endotoxic shock. Cell. 2004;116(4):617–628. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
