

Detergent-free isolation, characterization, and functional reconstitution of a tetrameric K+ channel: the power of native nanodiscs
- PMID: 25512535
- PMCID: PMC4284610
- DOI: 10.1073/pnas.1416205112
Detergent-free isolation, characterization, and functional reconstitution of a tetrameric K+ channel: the power of native nanodiscs
Abstract
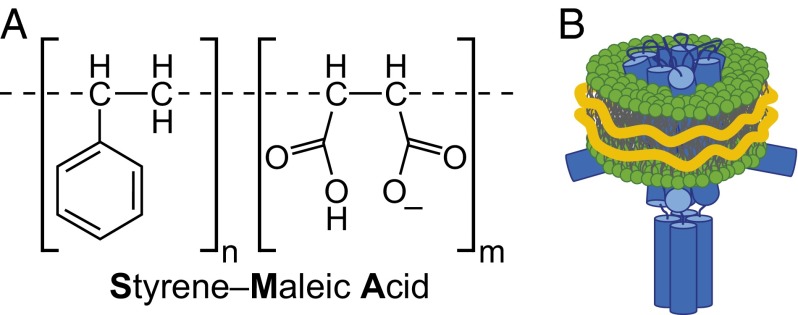
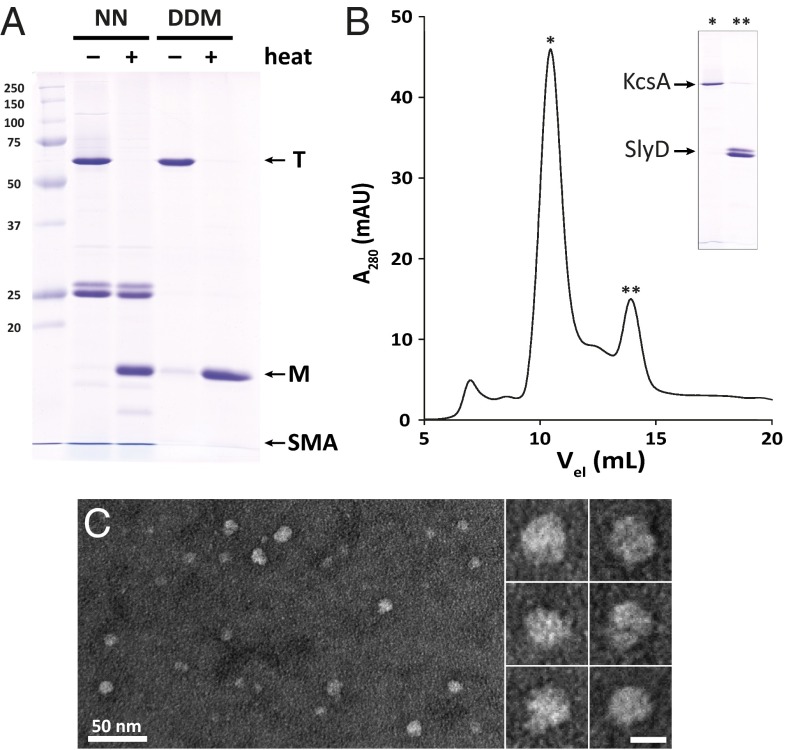
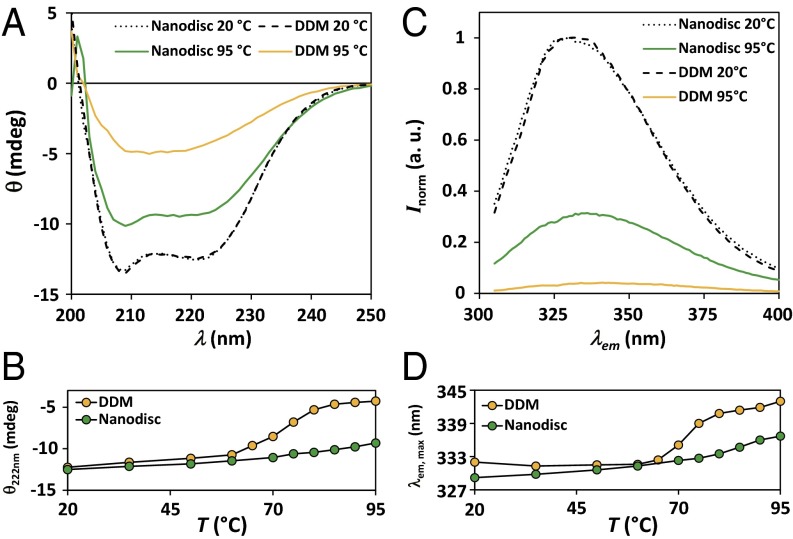
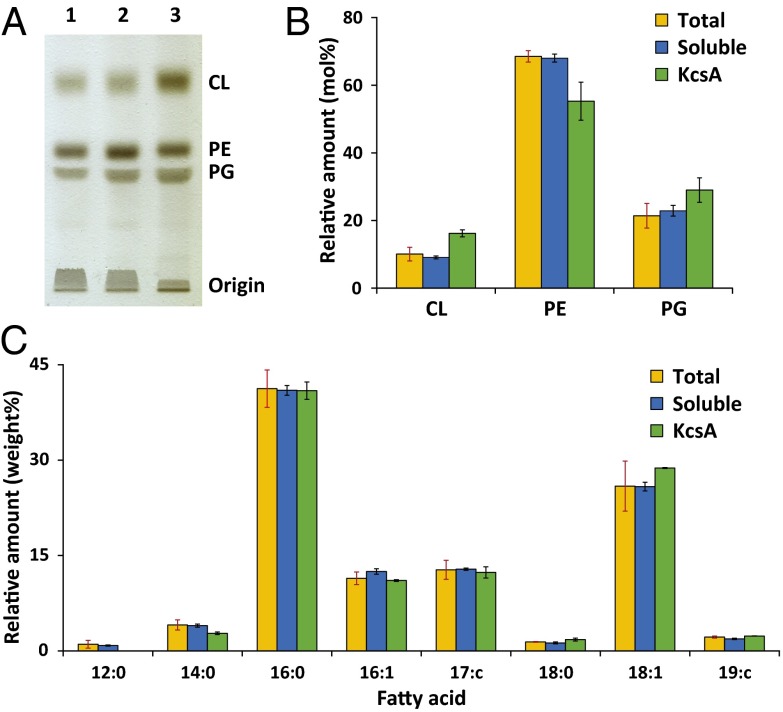
A major obstacle in the study of membrane proteins is their solubilization in a stable and active conformation when using detergents. Here, we explored a detergent-free approach to isolating the tetrameric potassium channel KcsA directly from the membrane of Escherichia coli, using a styrene-maleic acid copolymer. This polymer self-inserts into membranes and is capable of extracting membrane patches in the form of nanosize discoidal proteolipid particles or "native nanodiscs." Using circular dichroism and tryptophan fluorescence spectroscopy, we show that the conformation of KcsA in native nanodiscs is very similar to that in detergent micelles, but that the thermal stability of the protein is higher in the nanodiscs. Furthermore, as a promising new application, we show that quantitative analysis of the co-isolated lipids in purified KcsA-containing nanodiscs allows determination of preferential lipid-protein interactions. Thin-layer chromatography experiments revealed an enrichment of the anionic lipids cardiolipin and phosphatidylglycerol, indicating their close proximity to the channel in biological membranes and supporting their functional relevance. Finally, we demonstrate that KcsA can be reconstituted into planar lipid bilayers directly from native nanodiscs, which enables functional characterization of the channel by electrophysiology without first depriving the protein of its native environment. Together, these findings highlight the potential of the use of native nanodiscs as a tool in the study of ion channels, and of membrane proteins in general.
Keywords: ion channels; lipid–protein interactions; membrane–protein solubilization; nanodisc; styrene-maleic acid copolymer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- von Heijne G. The membrane protein universe: What’s out there and why bother? J Intern Med. 2007;261(6):543–557. - PubMed
-
- Garavito RM, Ferguson-Miller S. Detergents as tools in membrane biochemistry. J Biol Chem. 2001;276(35):32403–32406. - PubMed
-
- McQuade DT, et al. Rigid amphiphiles for membrane protein manipulation. Angew Chem Int Ed Engl. 2000;39(4):758–761. - PubMed
-
- Bowie JU. Stabilizing membrane proteins. Curr Opin Struct Biol. 2001;11(4):397–402. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
