

Plasminogen activation by staphylokinase enhances local spreading of S. aureus in skin infections
- PMID: 25515118
- PMCID: PMC4274676
- DOI: 10.1186/s12866-014-0310-7
Plasminogen activation by staphylokinase enhances local spreading of S. aureus in skin infections
Abstract
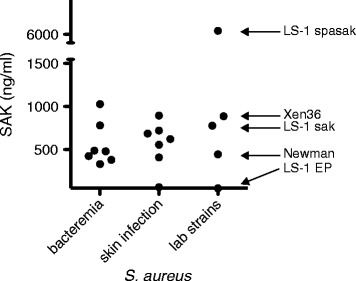
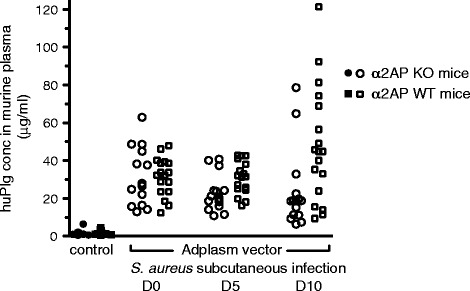
Background: Staphylococcus aureus (S. aureus) is a frequent cause of skin and soft tissue infections. A unique feature of S. aureus is the combined presence of coagulases that trigger fibrin formation and of the plasminogen activator staphylokinase (SAK). Whereas the importance of fibrin generation for S. aureus virulence has been established, the role of SAK remains unclear. We studied the role of plasminogen activation by SAK in a skin infection model in mice and evaluated the impact of alpha-2-antiplasmin (α2AP) deficiency on the spreading and proteolytic activity of S. aureus skin infections. The species-selectivity of SAK was overcome by adenoviral expression of human plasminogen. Bacterial spread and density was assessed non-invasively by imaging the bioluminescence of S. aureus Xen36.
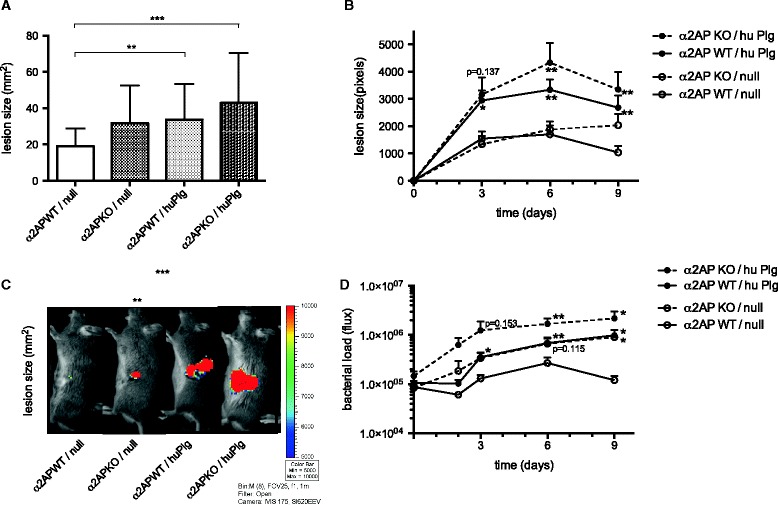
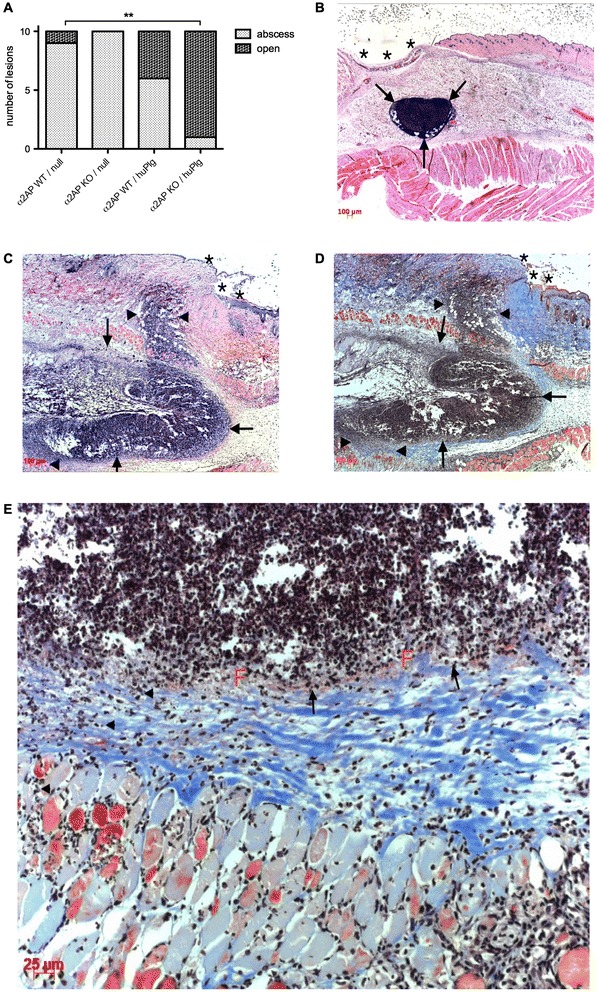
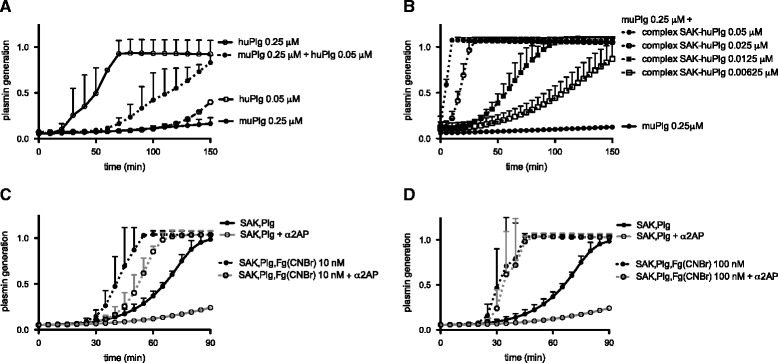
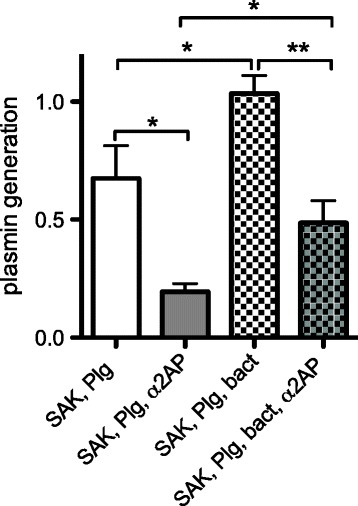
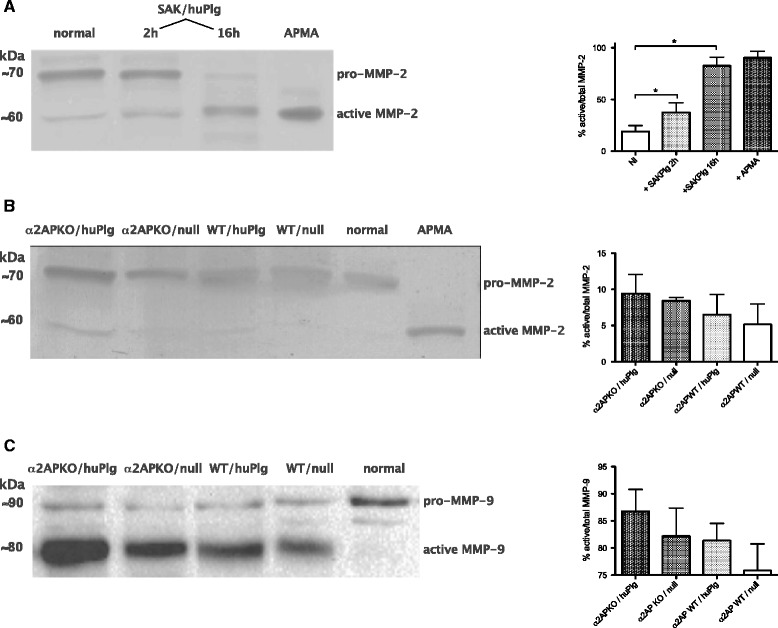
Results: SAK-mediated plasmin activity increased the local invasiveness of S. aureus, leading to larger lesions with skin disruption as well as decreased bacterial clearance by the host. Even though fibrin and bacterial surfaces protected SAK-mediated plasmin activity from inhibition by α2AP, the deficiency of α2AP resulted in increased bacterial spreading. SAK-mediated plasmin also induced secondary activation of gelatinases, shown both in vitro and in lesions from the in vivo model.
Conclusion: SAK contributes to the phenotype of S. aureus skin infections by enhancing bacterial spreading as a result of fibrinolytic and proteolytic activation.
Figures
References
-
- Vanassche T, Peetermans M, Van Aelst L, Peetermans WE, Verhaegen J, Missiakas D, Schneewind O, Hoylaerts M, Verhamme P. The role of staphylothrombin-mediated fibrin deposition in catheter-related Staphylococcus aureus infections. J Infect Dis. 2013;208:92–100. doi: 10.1093/infdis/jit130. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
