

A large animal model of spinal muscular atrophy and correction of phenotype
- PMID: 25516063
- PMCID: PMC4453930
- DOI: 10.1002/ana.24332
A large animal model of spinal muscular atrophy and correction of phenotype
Abstract
Objectives: Spinal muscular atrophy (SMA) is caused by reduced levels of survival motor neuron (SMN) protein, which results in motoneuron loss. Therapeutic strategies to increase SMN levels including drug compounds, antisense oligonucleotides, and scAAV9 gene therapy have proved effective in mice. We wished to determine whether reduction of SMN in postnatal motoneurons resulted in SMA in a large animal model, whether SMA could be corrected after development of muscle weakness, and the response of clinically relevant biomarkers.
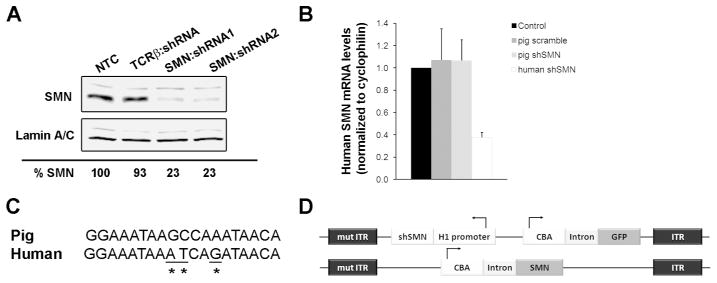
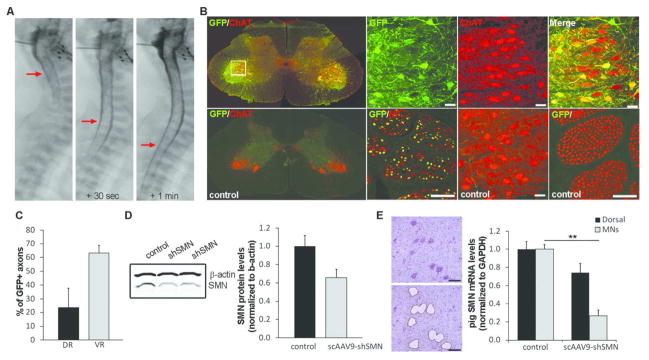
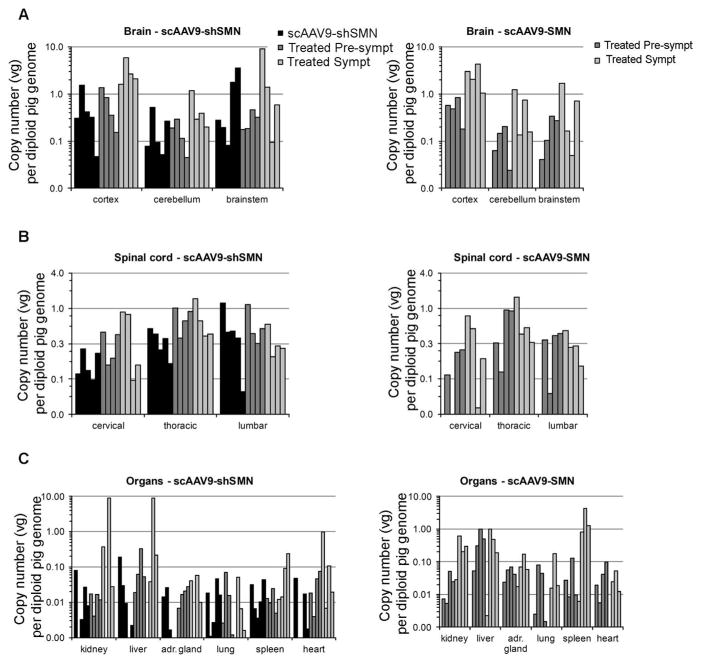
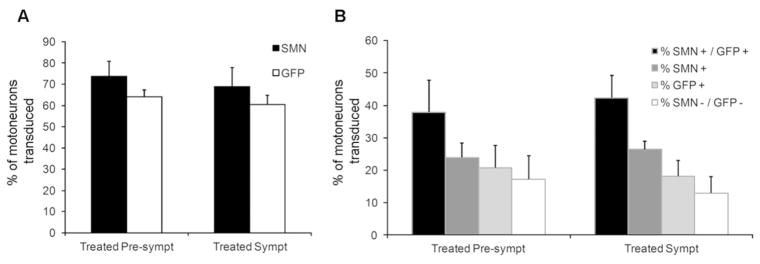
Methods: Using intrathecal delivery of scAAV9 expressing an shRNA targeting pig SMN1, SMN was knocked down in motoneurons postnatally to SMA levels. This resulted in an SMA phenotype representing the first large animal model of SMA. Restoration of SMN was performed at different time points with scAAV9 expressing human SMN (scAAV9-SMN), and electrophysiology measurements and pathology were performed.
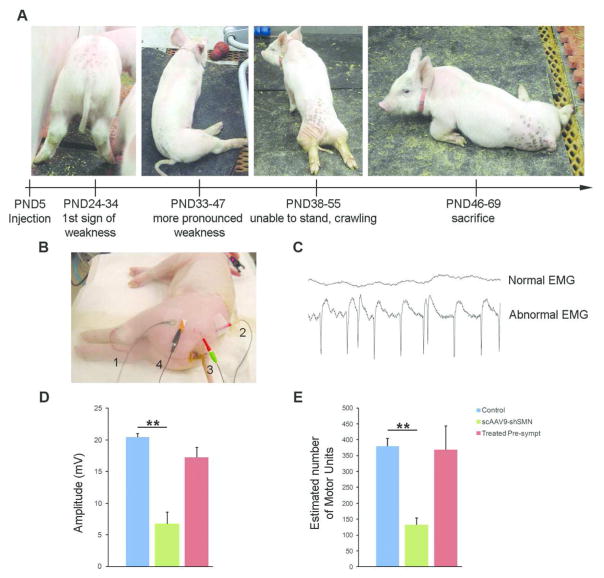
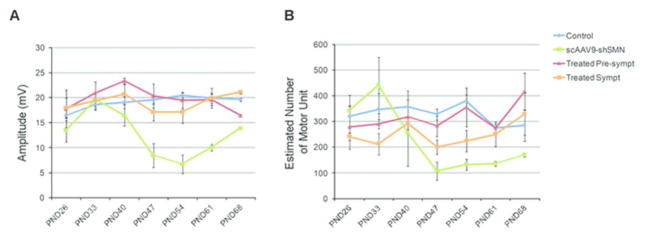
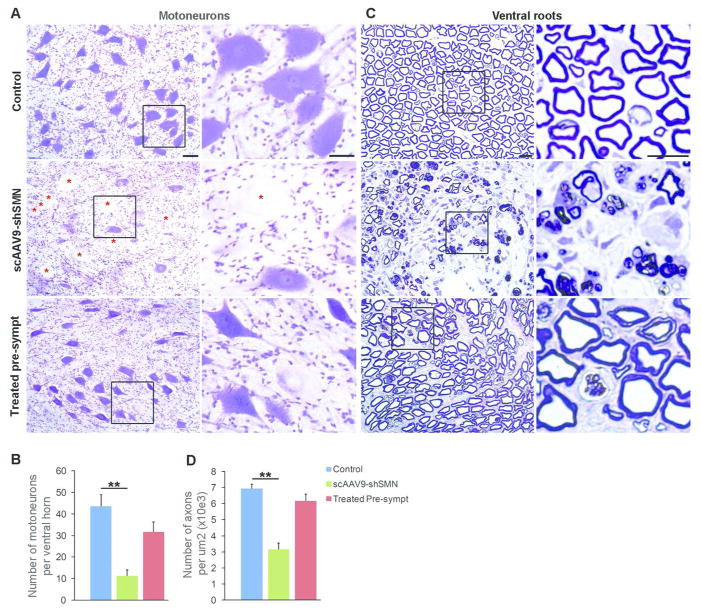
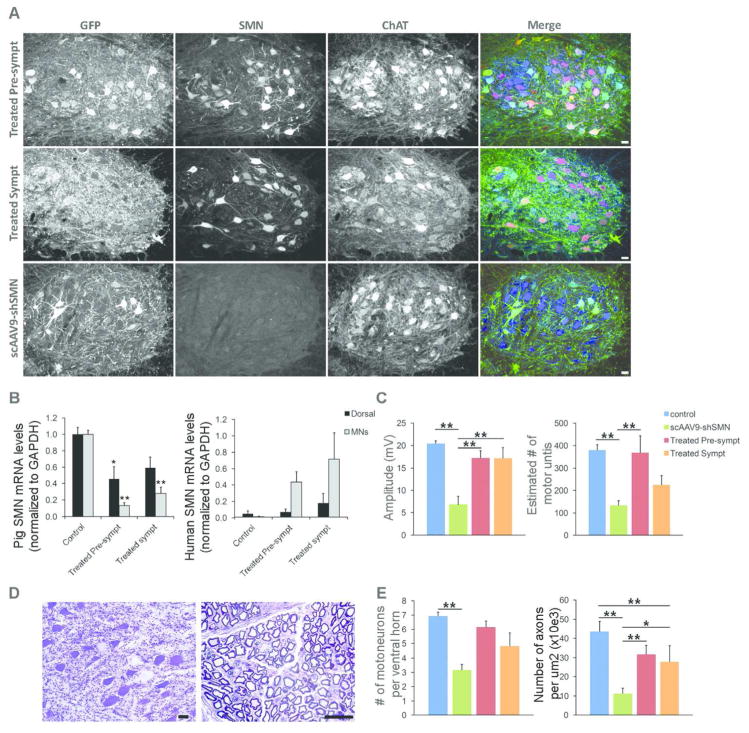
Results: Knockdown of SMN in postnatal motoneurons results in overt proximal weakness, fibrillations on electromyography indicating active denervation, and reduced compound muscle action potential (CMAP) and motor unit number estimation (MUNE), as in human SMA. Neuropathology showed loss of motoneurons and motor axons. Presymptomatic delivery of scAAV9-SMN prevented SMA symptoms, indicating that all changes are SMN dependent. Delivery of scAAV9-SMN after symptom onset had a marked impact on phenotype, electrophysiological measures, and pathology.
Interpretation: High SMN levels are critical in postnatal motoneurons, and reduction of SMN results in an SMA phenotype that is SMN dependent. Importantly, clinically relevant biomarkers including CMAP and MUNE are responsive to SMN restoration, and abrogation of phenotype can be achieved even after symptom onset.
© 2014 American Neurological Association.
Conflict of interest statement
Figures
References
-
- Prior TW, Snyder PJ, Rink BD, et al. Newborn and carrier screening for spinal muscular atrophy. Am J Med Genet A. 2010;152A7:1608–1616. - PubMed
-
- Monani UR, Lorson CL, Parsons DW, et al. A single nucleotide difference that alters splicing patterns distinguishes the SMA gene SMN1 from the copy gene SMN2. Hum Mol Genet. 1999;87:1177–1183. - PubMed
-
- Lefebvre S, Burglen L, Reboullet S, et al. Identification and characterization of a spinal muscular atrophy-determining gene. Cell. 1995;801:155–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
