

Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions
- PMID: 25516826
- PMCID: PMC4266812
- DOI: 10.1016/j.ijpddr.2014.07.007
Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions
Abstract
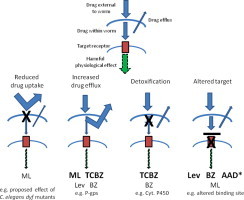
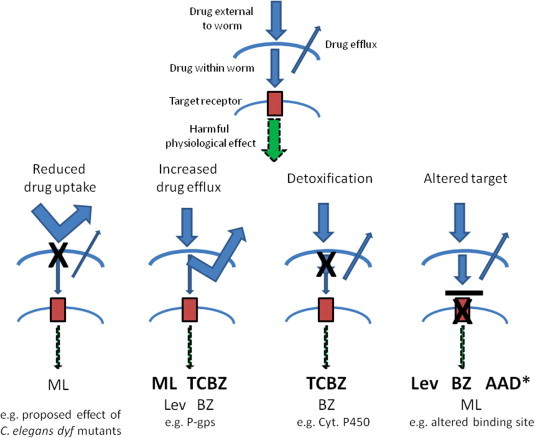
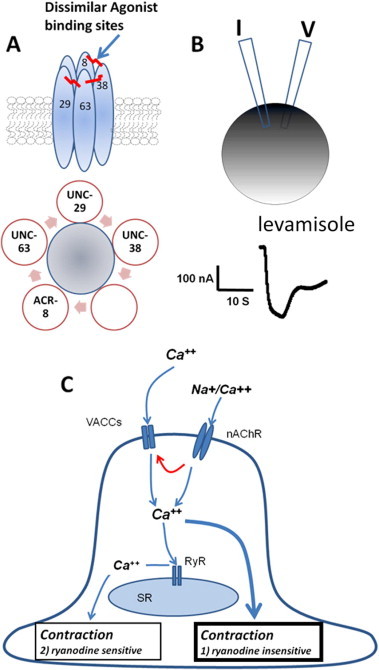
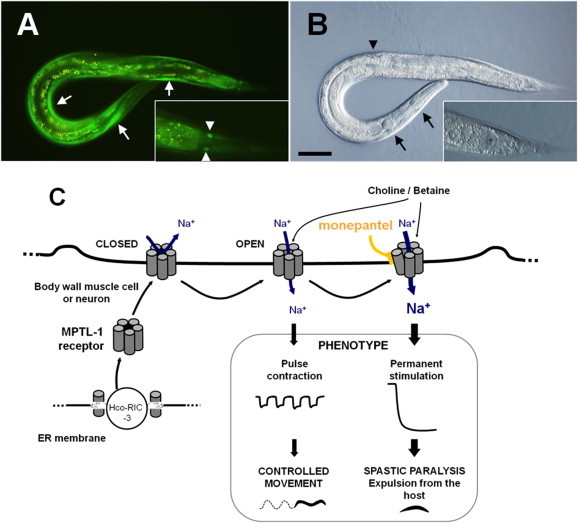
Anthelmintic resistance has a great impact on livestock production systems worldwide, is an emerging concern in companion animal medicine, and represents a threat to our ongoing ability to control human soil-transmitted helminths. The Consortium for Anthelmintic Resistance and Susceptibility (CARS) provides a forum for scientists to meet and discuss the latest developments in the search for molecular markers of anthelmintic resistance. Such markers are important for detecting drug resistant worm populations, and indicating the likely impact of the resistance on drug efficacy. The molecular basis of resistance is also important for understanding how anthelmintics work, and how drug resistant populations arise. Changes to target receptors, drug efflux and other biological processes can be involved. This paper reports on the CARS group meeting held in August 2013 in Perth, Australia. The latest knowledge on the development of molecular markers for resistance to each of the principal classes of anthelmintics is reviewed. The molecular basis of resistance is best understood for the benzimidazole group of compounds, and we examine recent work to translate this knowledge into useful diagnostics for field use. We examine recent candidate-gene and whole-genome approaches to understanding anthelmintic resistance and identify markers. We also look at drug transporters in terms of providing both useful markers for resistance, as well as opportunities to overcome resistance through the targeting of the transporters themselves with inhibitors. Finally, we describe the tools available for the application of the newest high-throughput sequencing technologies to the study of anthelmintic resistance.
Keywords: Anthelmintic drugs; Anthelmintic resistance; Anthelmintic targets; Molecular markers; Receptors.
Figures

References
-
- Abad P., Gouzy J., Aury J.M., Castagnone-Sereno P., Danchin E.G., Deleury E., Perfus-Barbeoch L., Anthouard V., Artiguenave F., Blok V.C., Caillaud M.C., Coutinho P.M., Dasilva C., De Luca F., Deau F., Esquibet M., Flutre T., Goldstone J.V., Hamamouch N., Hewezi T., Jaillon O., Jubin C., Leonetti P., Magliano M., Maier T.R., Markov G.V., McVeigh P., Pesole G., Poulain J., Robinson-Rechavi M., Sallet E., Segurens B., Steinbach D., Tytgat T., Ugarte E., van Ghelder C., Veronico P., Baum T.J., Blaxter M., Bleve-Zacheo T., Davis E.L., Ewbank J.J., Favery B., Grenier E., Henrissat B., Jones J.T., Laudet V., Maule A.G., Quesneville H., Rosso M.N., Schiex T., Smant G., Weissenbach J., Wincker P. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008;26:909–915. - PubMed
-
- Adelsberger H., Lepier A., Dudel J. Activation of rat recombinant alpha(1)beta(2)gamma(2S) GABA(A) receptor by the insecticide ivermectin. Eur. J. Pharmacol. 2000;394:163–170. - PubMed
-
- Alvarez L.L., Solana H.D., Mottier M.L., Virkel G.L., Fairweather I., Lanusse C.E. Altered drug influx/efflux and enhanced metabolic activity in triclabendazole-resistant liver flukes. Parasitology. 2005;131:501–510. - PubMed
-
- Ardelli B.F., Prichard R.K. Reduced genetic variation of an Onchocerca volvulus ABC transporter gene following treatment with ivermectin. Trans. R. Soc. Trop. Med. Hyg. 2007;101:1223–1232. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
