

Dauer-independent insulin/IGF-1-signalling implicates collagen remodelling in longevity
- PMID: 25517099
- PMCID: PMC4352135
- DOI: 10.1038/nature14021
Dauer-independent insulin/IGF-1-signalling implicates collagen remodelling in longevity
Abstract
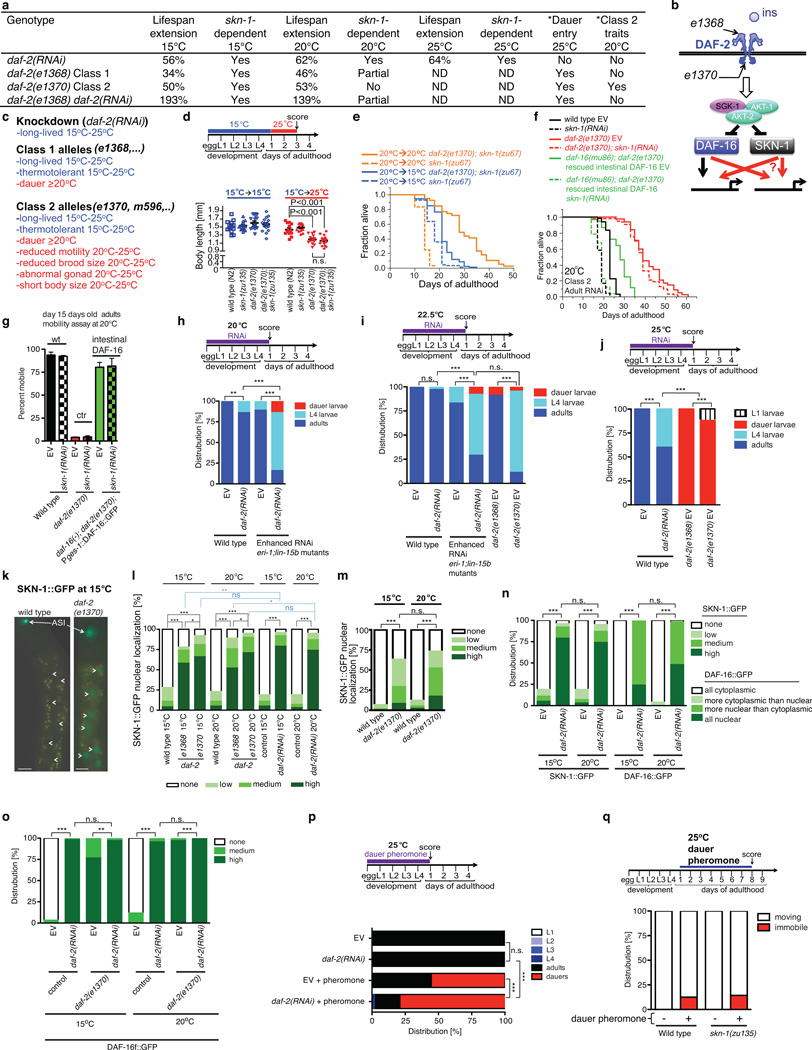
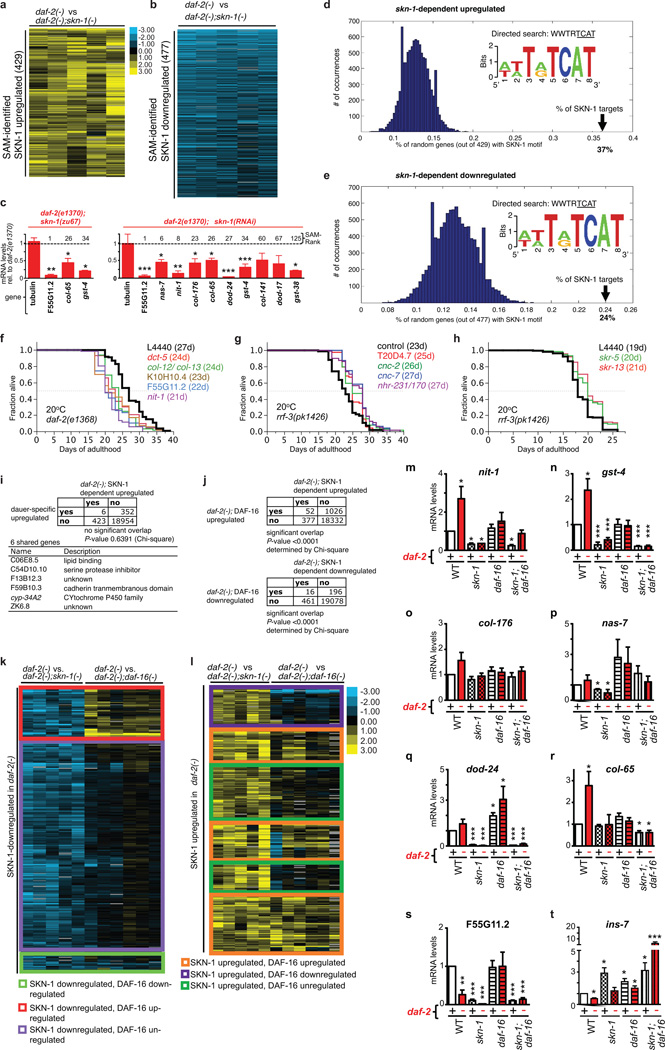
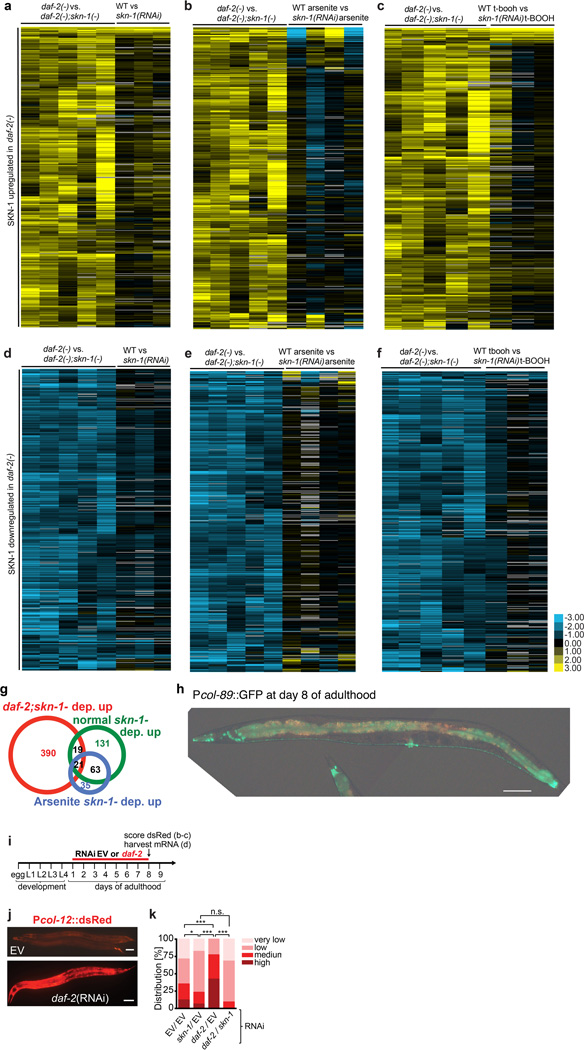
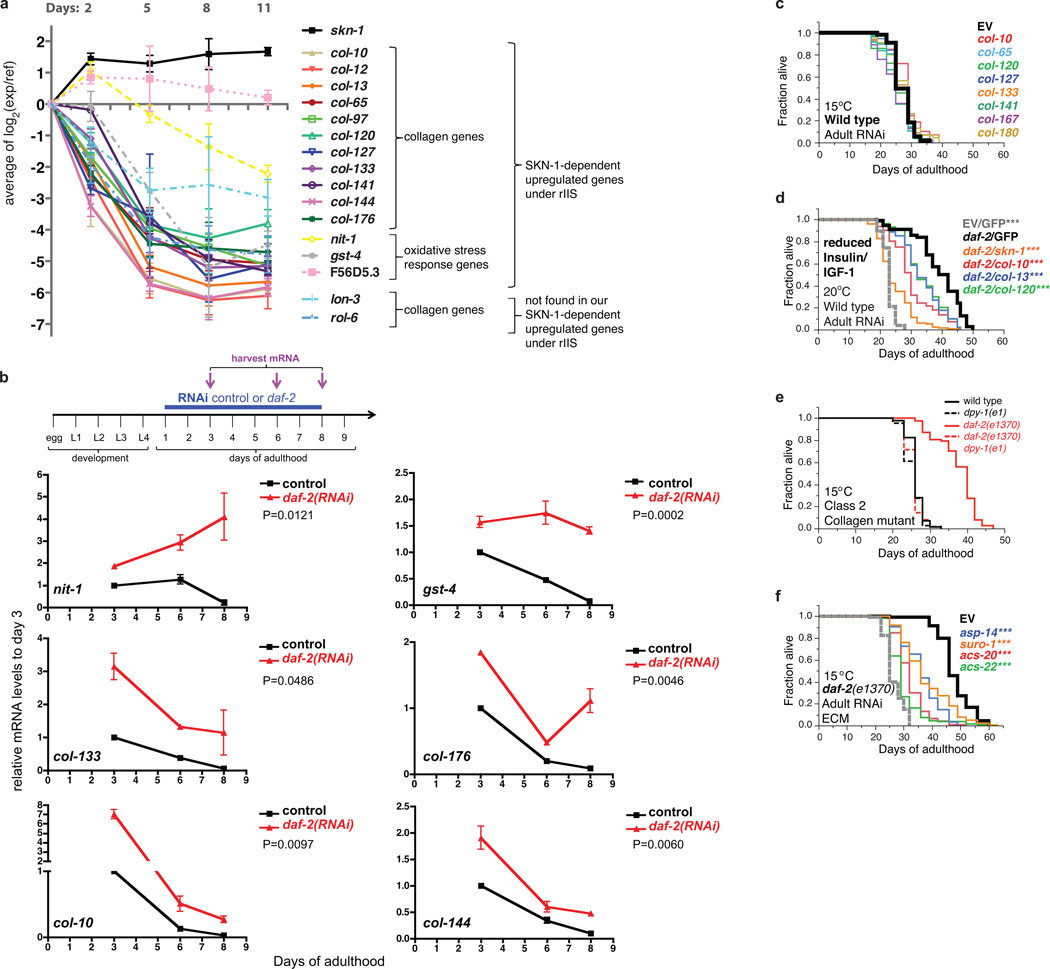
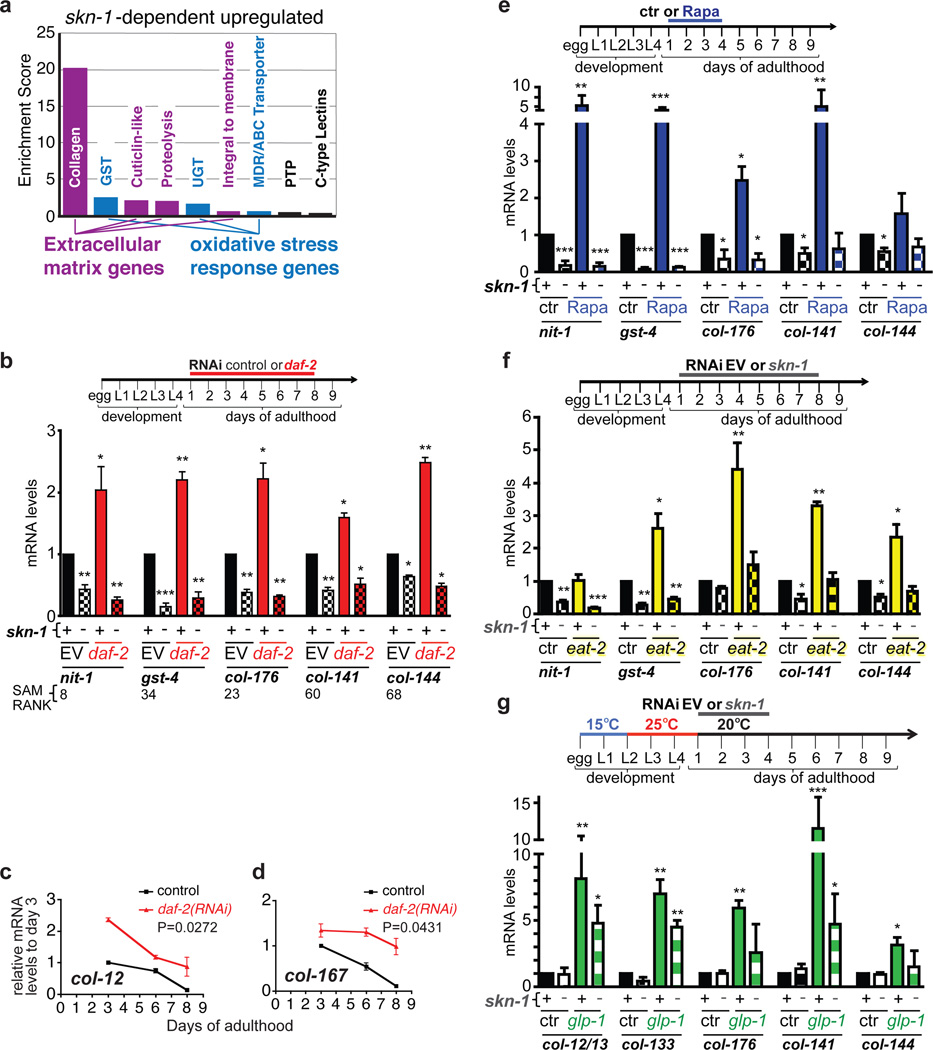
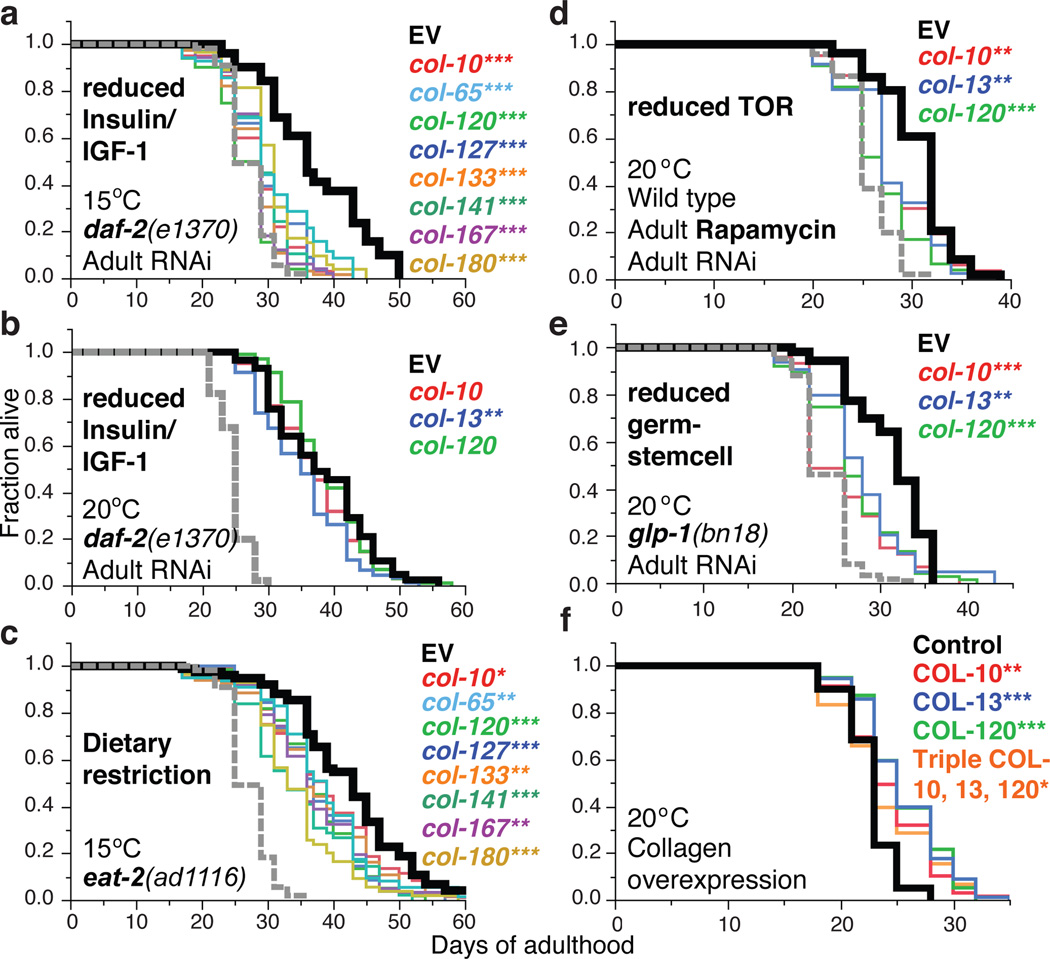
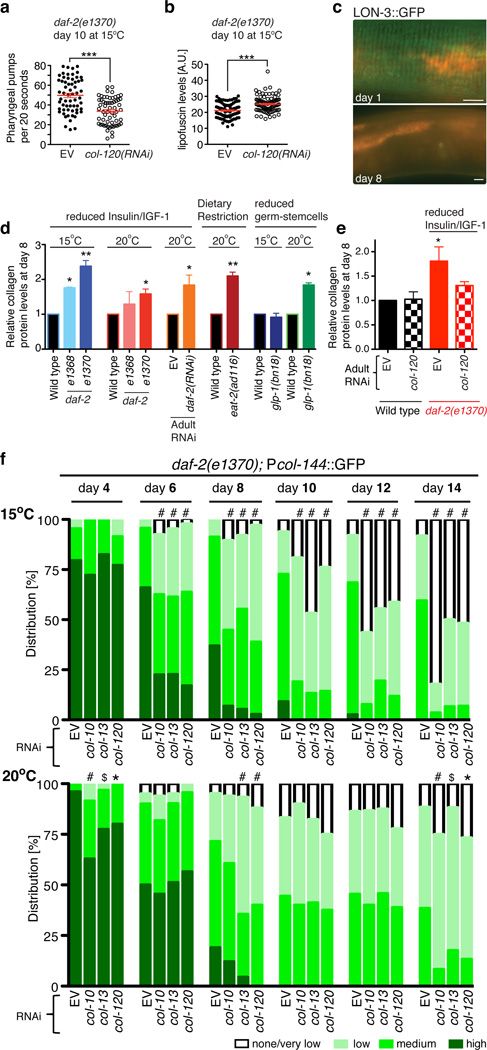
Interventions that delay ageing mobilize mechanisms that protect and repair cellular components, but it is unknown how these interventions might slow the functional decline of extracellular matrices, which are also damaged during ageing. Reduced insulin/IGF-1 signalling (rIIS) extends lifespan across the evolutionary spectrum, and in juvenile Caenorhabditis elegans also allows the transcription factor DAF-16/FOXO to induce development into dauer, a diapause that withstands harsh conditions. It has been suggested that rIIS delays C. elegans ageing through activation of dauer-related processes during adulthood, but some rIIS conditions confer robust lifespan extension unaccompanied by any dauer-like traits. Here we show that rIIS can promote C. elegans longevity through a program that is genetically distinct from the dauer pathway, and requires the Nrf (NF-E2-related factor) orthologue SKN-1 acting in parallel to DAF-16. SKN-1 is inhibited by IIS and has been broadly implicated in longevity, but is rendered dispensable for rIIS lifespan extension by even mild activity of dauer-related processes. When IIS is decreased under conditions that do not induce dauer traits, SKN-1 most prominently increases expression of collagens and other extracellular matrix genes. Diverse genetic, nutritional, and pharmacological pro-longevity interventions delay an age-related decline in collagen expression. These collagens mediate adulthood extracellular matrix remodelling, and are needed for ageing to be delayed by interventions that do not involve dauer traits. By genetically delineating a dauer-independent rIIS ageing pathway, our results show that IIS controls a broad set of protective mechanisms during C. elegans adulthood, and may facilitate elucidation of processes of general importance for longevity. The importance of collagen production in diverse anti-ageing interventions implies that extracellular matrix remodelling is a generally essential signature of longevity assurance, and that agents promoting extracellular matrix youthfulness may have systemic benefit.
Figures
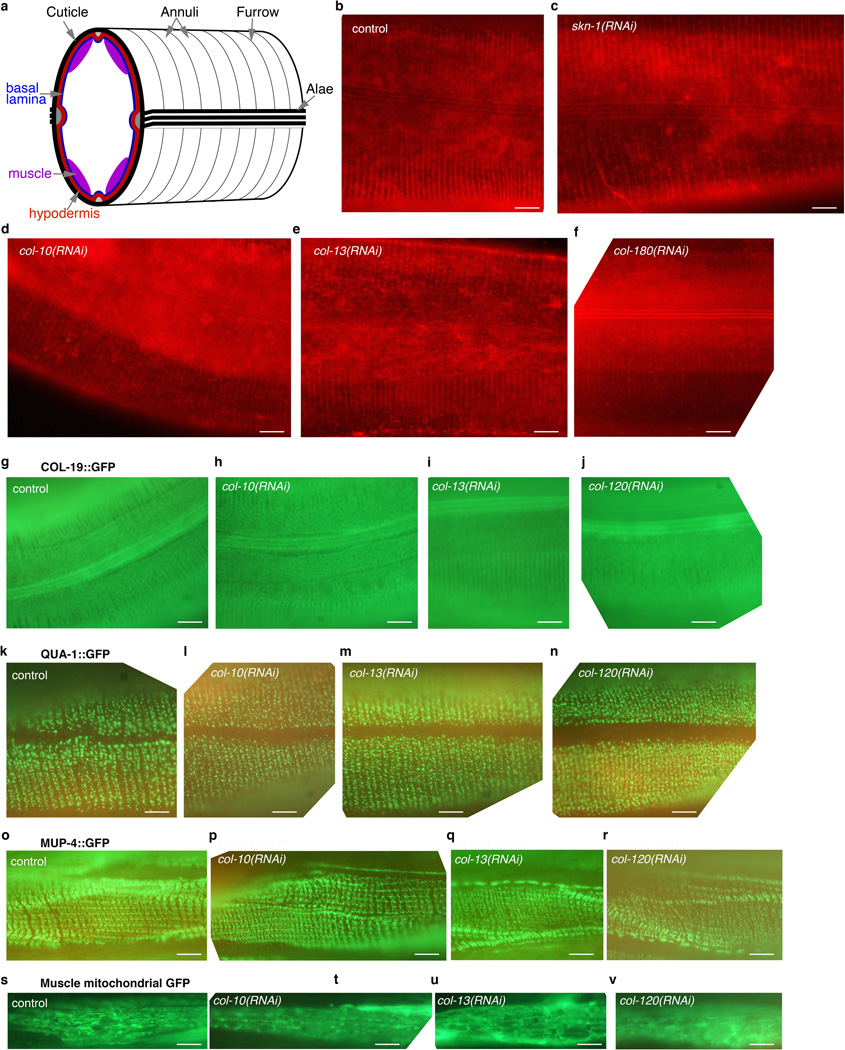
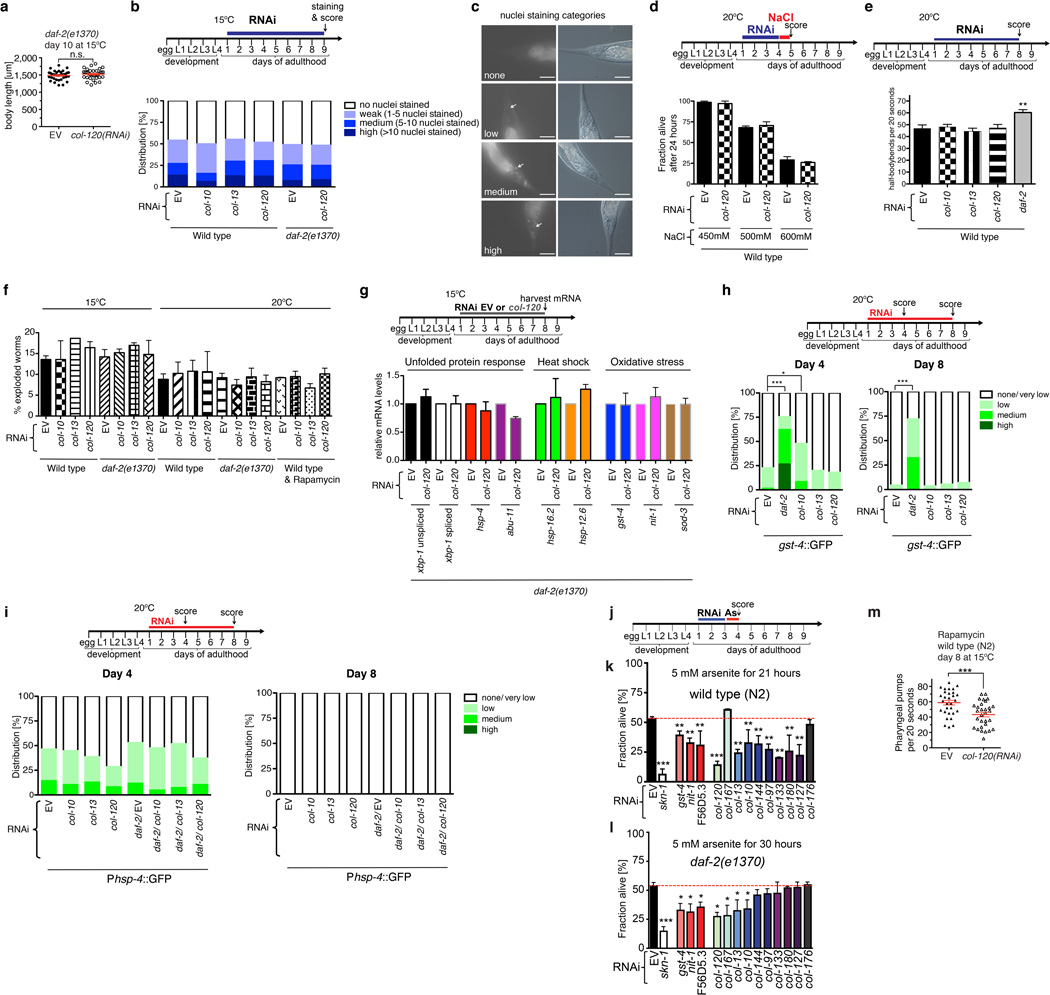
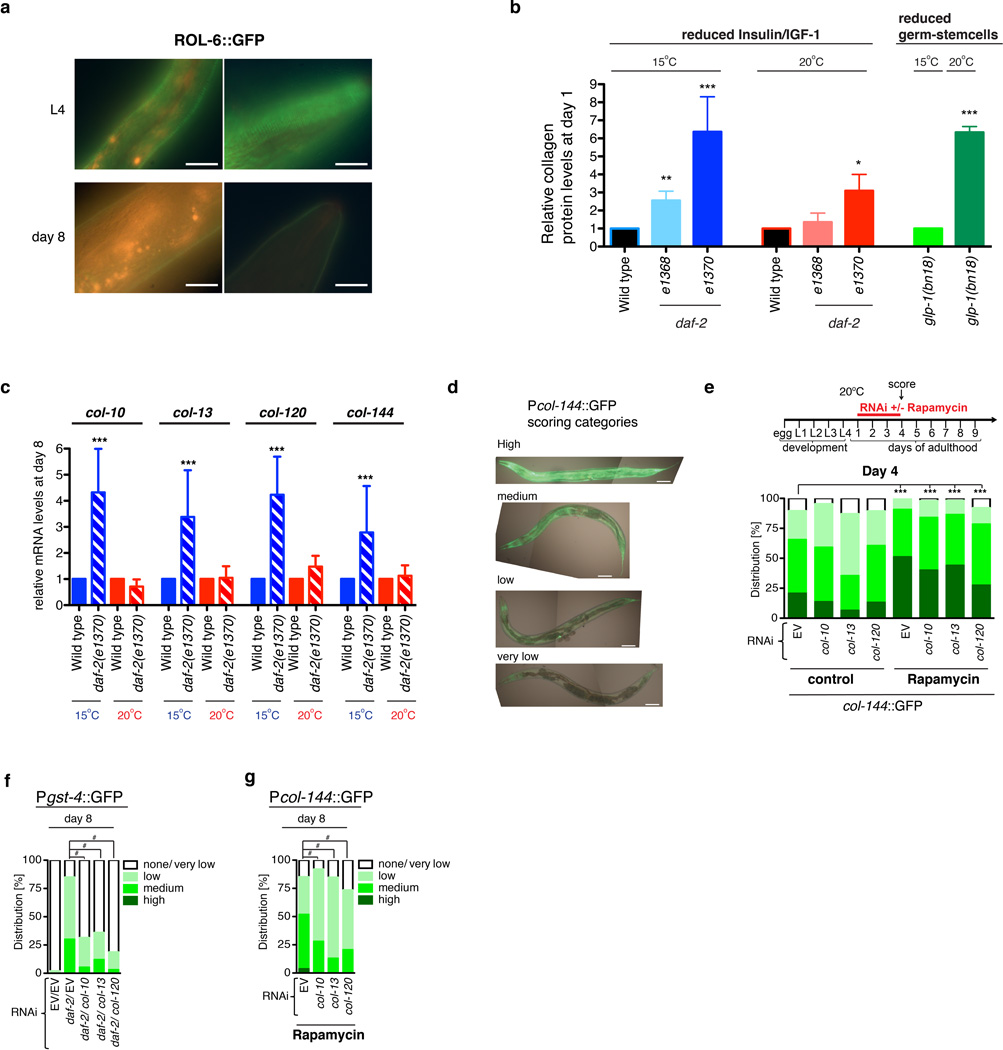
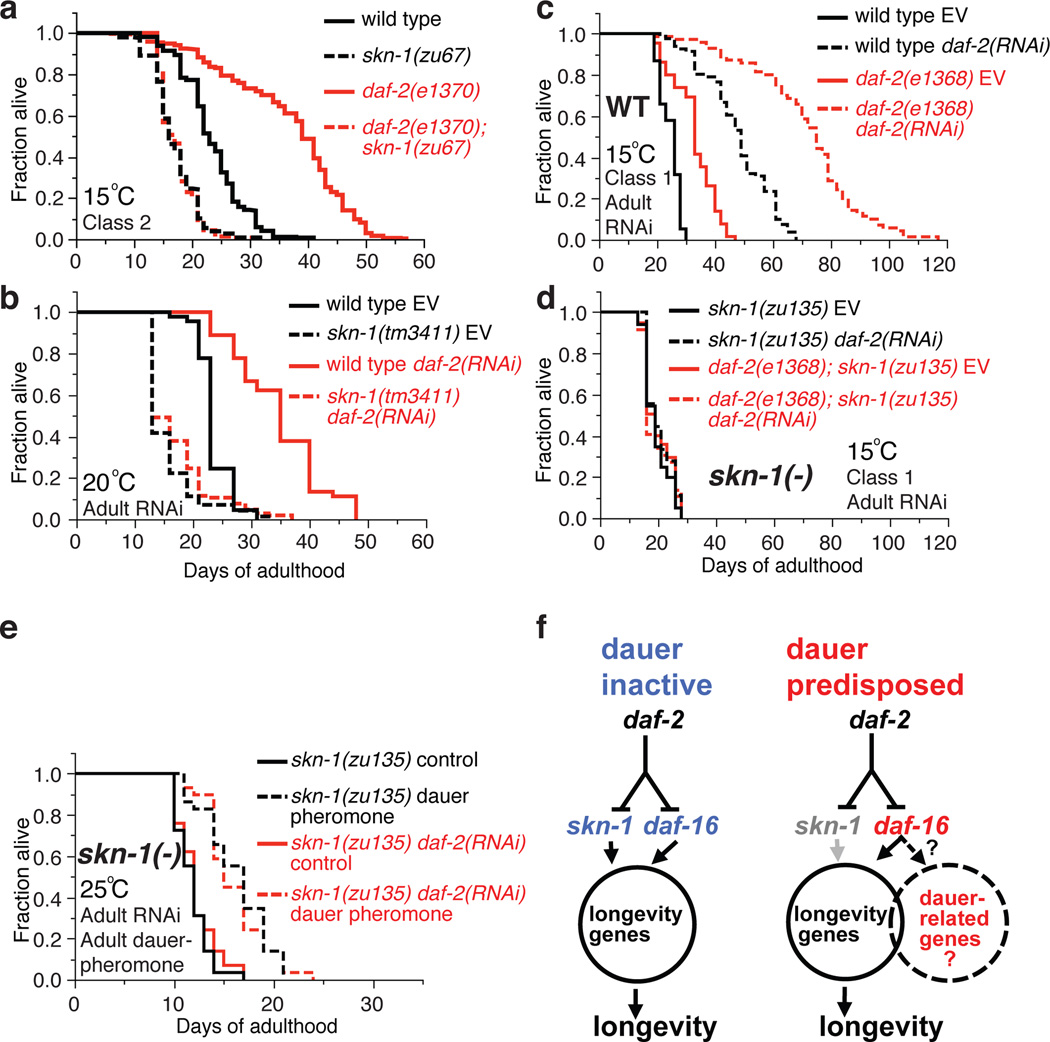
References
-
- Kenyon CJ. The genetics of ageing. Nature. 2010;464:504–512. doi:nature08980 [pii] 10.1038/nature08980. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
