

STING-dependent cytosolic DNA sensing mediates innate immune recognition of immunogenic tumors
- PMID: 25517615
- PMCID: PMC4384884
- DOI: 10.1016/j.immuni.2014.10.017
STING-dependent cytosolic DNA sensing mediates innate immune recognition of immunogenic tumors
Erratum in
- Immunity. 2015 Jan 20;42(1):199
Abstract
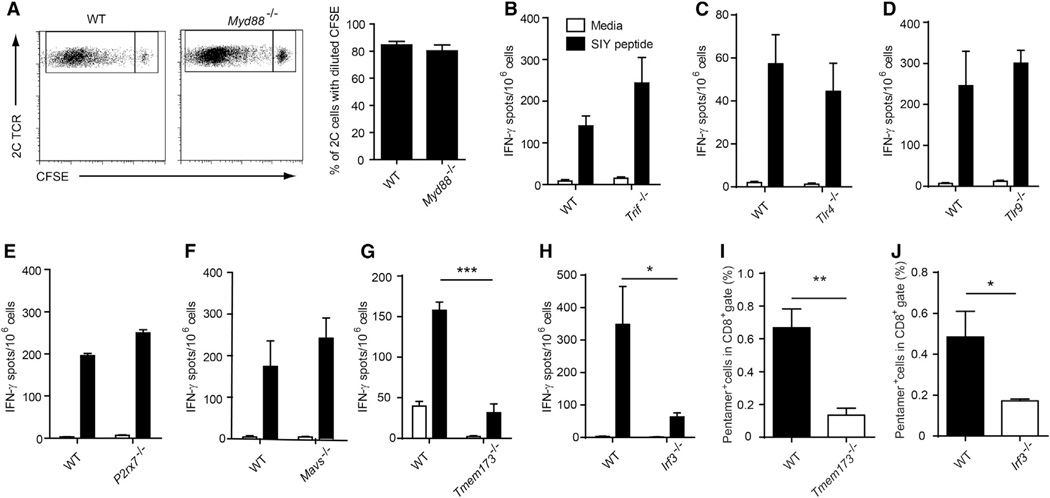
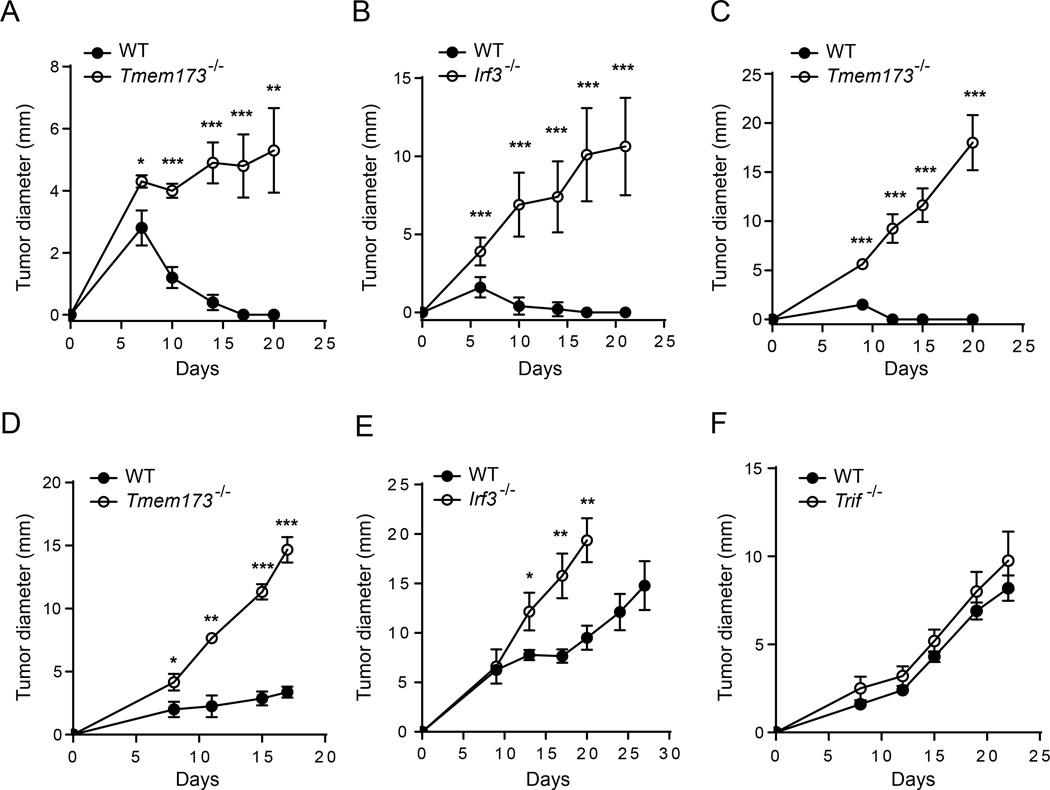
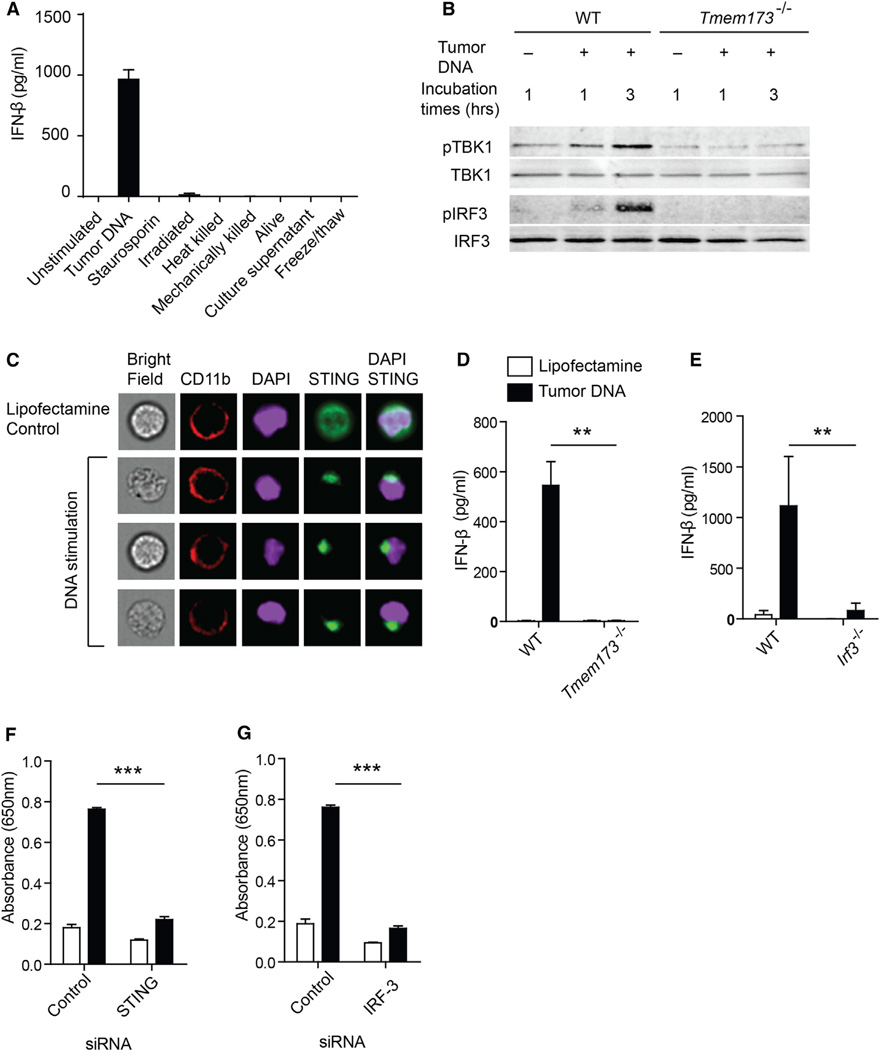
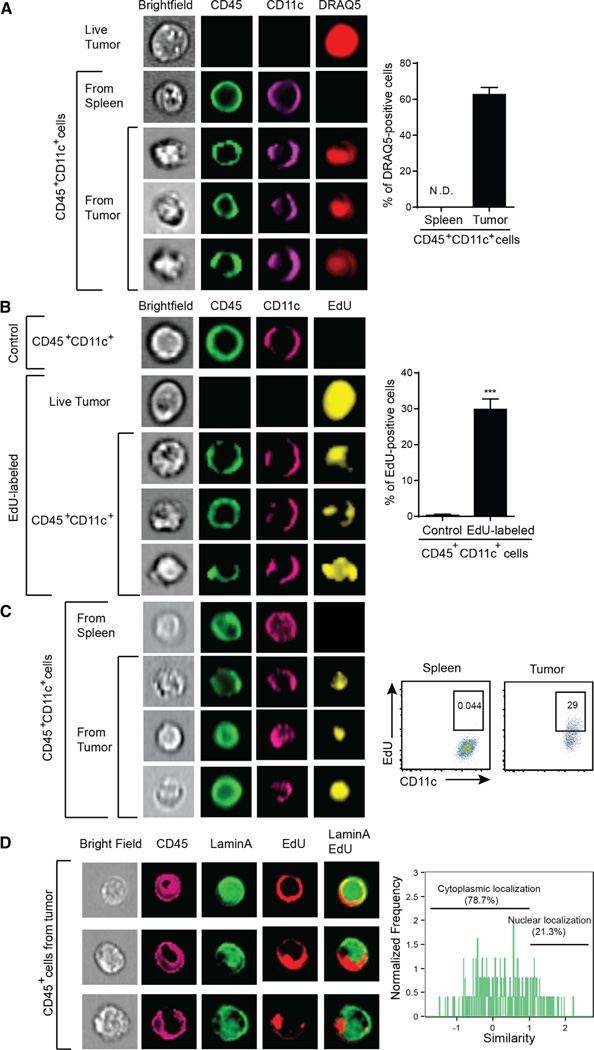
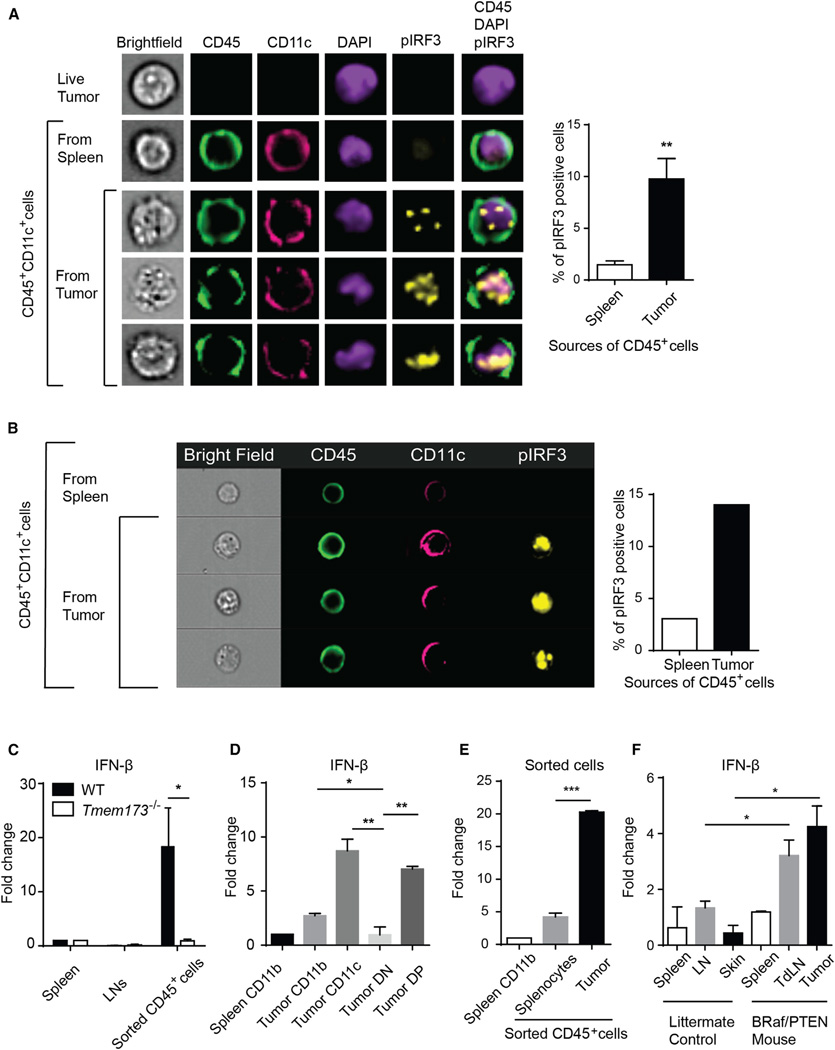
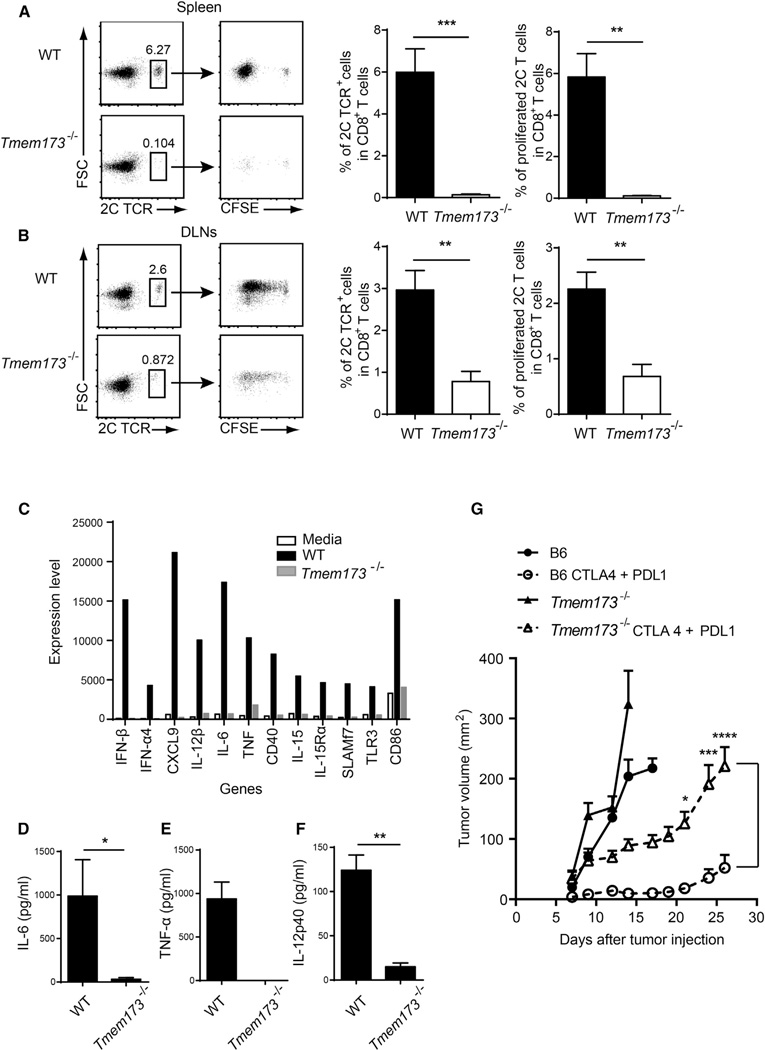
Spontaneous T cell responses against tumors occur frequently and have prognostic value in patients. The mechanism of innate immune sensing of immunogenic tumors leading to adaptive T cell responses remains undefined, although type I interferons (IFNs) are implicated in this process. We found that spontaneous CD8(+) T cell priming against tumors was defective in mice lacking stimulator of interferon genes complex (STING), but not other innate signaling pathways, suggesting involvement of a cytosolic DNA sensing pathway. In vitro, IFN-? production and dendritic cell activation were triggered by tumor-cell-derived DNA, via cyclic-GMP-AMP synthase (cGAS), STING, and interferon regulatory factor 3 (IRF3). In the tumor microenvironment in vivo, tumor cell DNA was detected within host antigen-presenting cells, which correlated with STING pathway activation and IFN-? production. Our results demonstrate that a major mechanism for innate immune sensing of cancer occurs via the host STING pathway, with major implications for cancer immunotherapy.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Tumors STING adaptive antitumor immunity.Immunity. 2014 Nov 20;41(5):679-81. doi: 10.1016/j.immuni.2014.11.004. Epub 2014 Nov 20. Immunity. 2014. PMID: 25517609
-
STINGing Antitumor Immunity into Action.Cancer Discov. 2018 Mar;8(3):259-260. doi: 10.1158/2159-8290.CD-ND2018-002. Epub 2018 Jan 25. Cancer Discov. 2018. PMID: 29371226
References
-
- Apetoh L, Ghiringhelli F, Tesniere A, Obeid M, Ortiz C, Criollo A, Mignot G, Maiuri MC, Ullrich E, Saulnier P, et al. Toll-like receptor 4-dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat. Med. 2007;13:1050–1059. - PubMed
-
- Blank C, Brown I, Peterson AC, Spiotto M, Iwai Y, Honjo T, Gajewski TF. PD-L1/B7H-1 inhibits the effector phase of tumor rejection by T cell receptor (TCR) transgenic CD8+ T cells. Cancer Res. 2004;64:1140–1145. - PubMed
-
- Blank C, Brown I, Kacha AK, Markiewicz MA, Gajewski TF. ICAM-1 contributes to but is not essential for tumor antigen cross-priming and CD8+ T cell-mediated tumor rejection in vivo. J. Immunol. 2005;174:3416–3420. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
