

Distinct epigenetic signatures delineate transcriptional programs during virus-specific CD8(+) T cell differentiation
- PMID: 25517617
- PMCID: PMC4479393
- DOI: 10.1016/j.immuni.2014.11.001
Distinct epigenetic signatures delineate transcriptional programs during virus-specific CD8(+) T cell differentiation
Erratum in
- Immunity. 2014 Dec 18;41(6):1064
Abstract
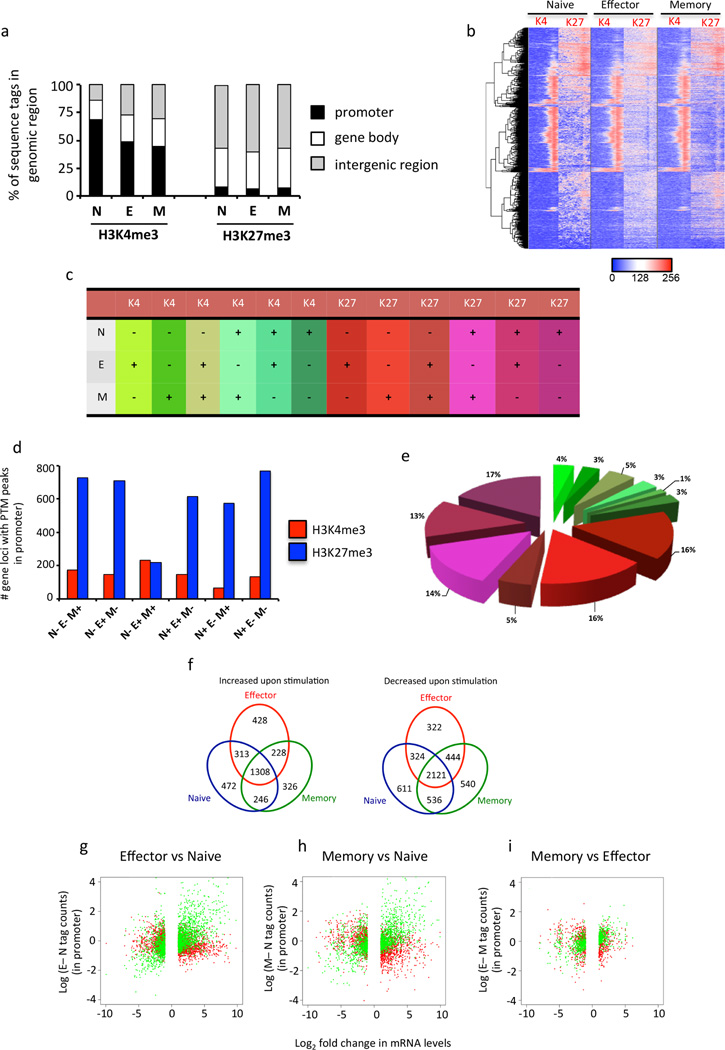
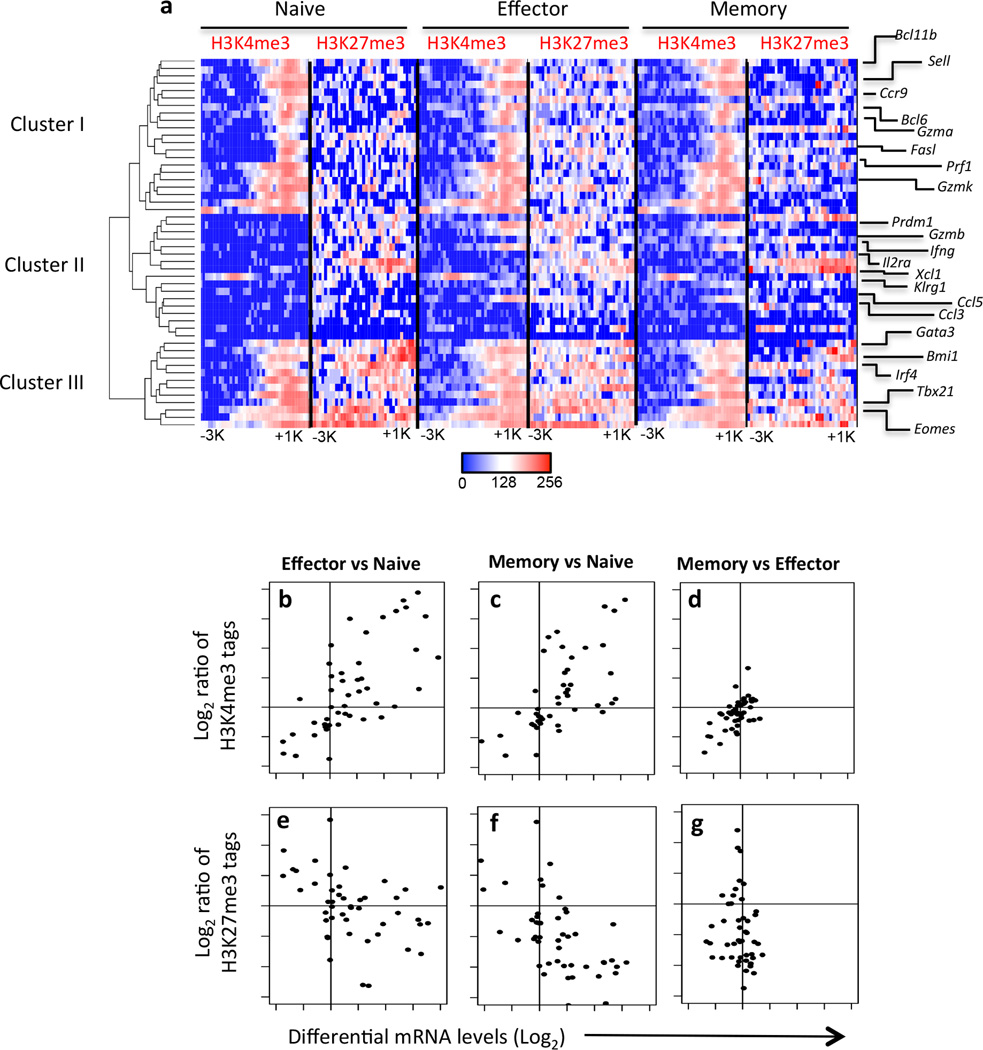
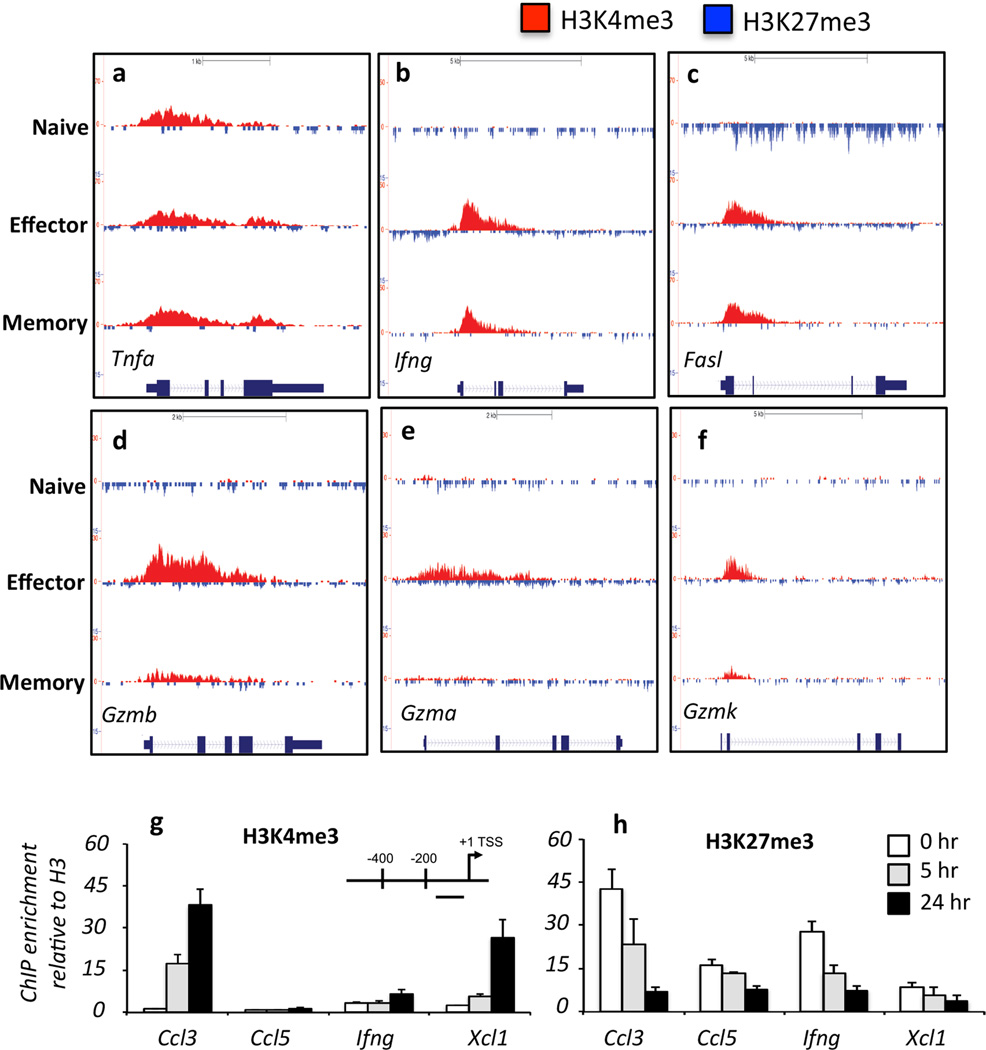
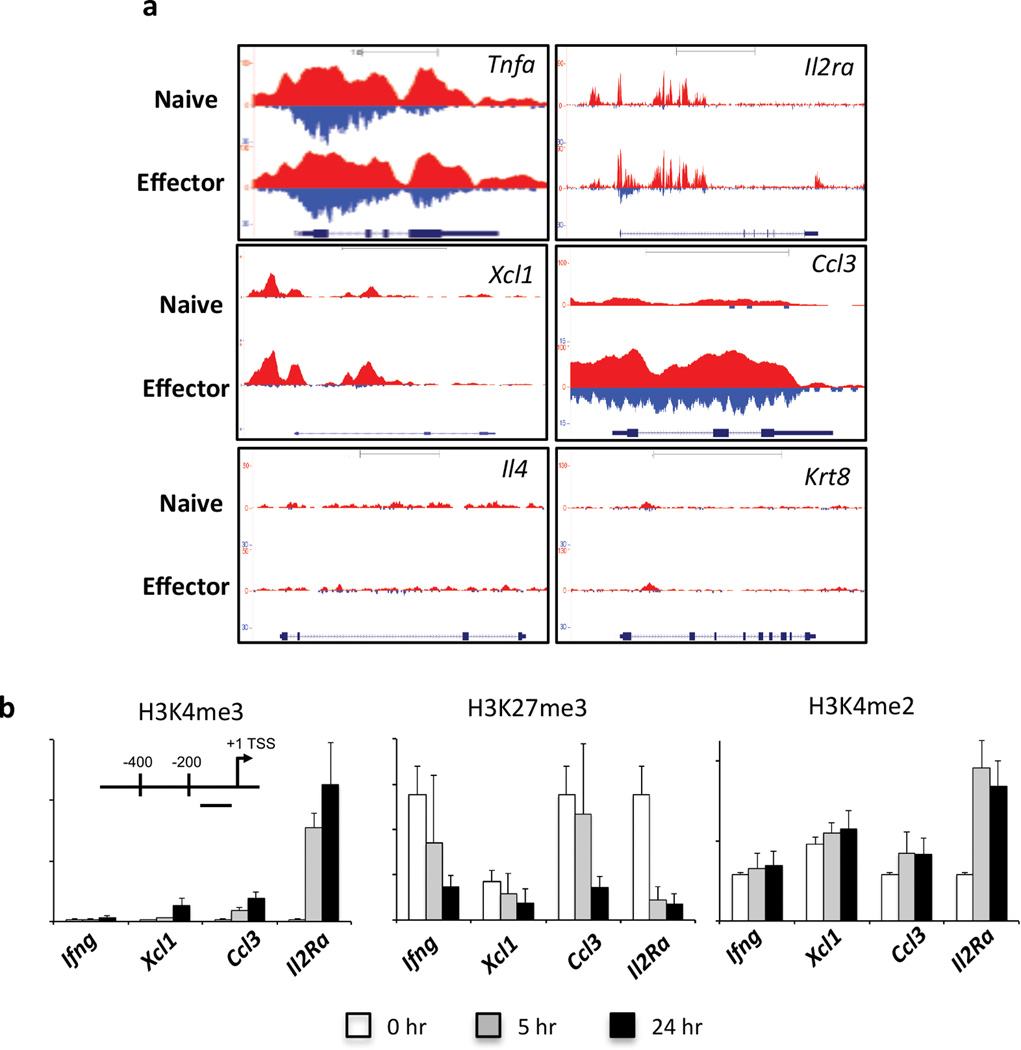
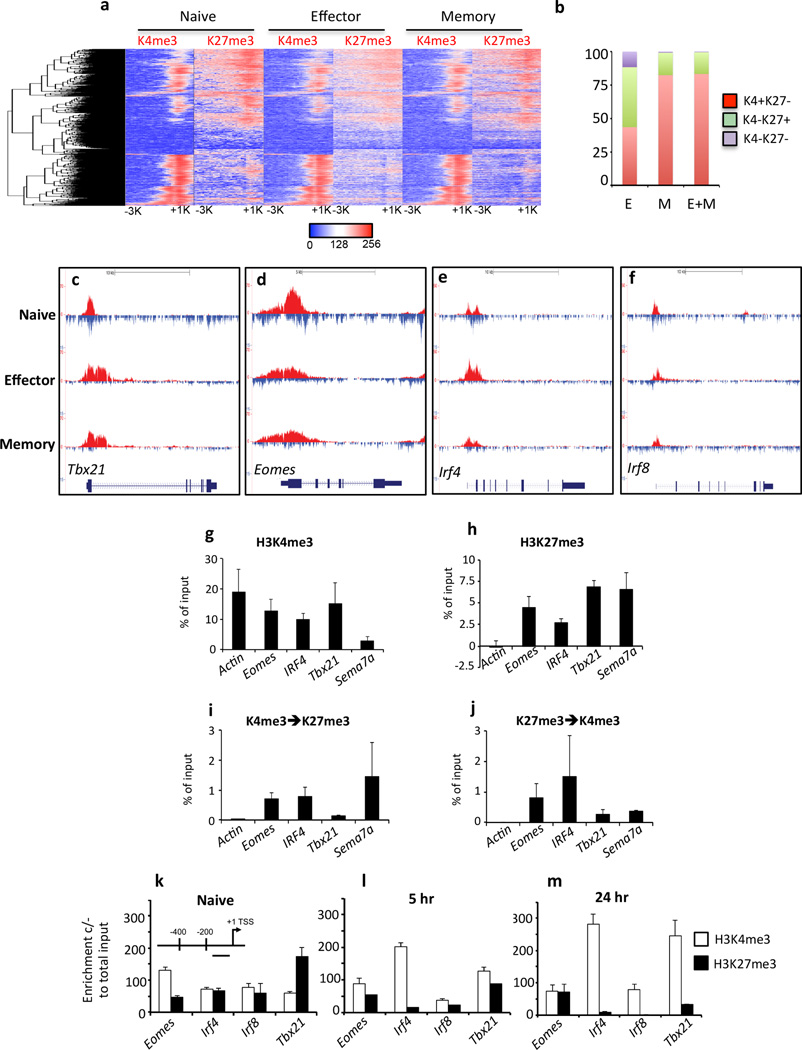
The molecular mechanisms that regulate the rapid transcriptional changes that occur during cytotoxic T lymphocyte (CTL) proliferation and differentiation in response to infection are poorly understood. We have utilized ChIP-seq to assess histone H3 methylation dynamics within naive, effector, and memory virus-specific T cells isolated directly ex vivo after influenza A virus infection. Our results show that within naive T cells, codeposition of the permissive H3K4me3 and repressive H3K27me3 modifications is a signature of gene loci associated with gene transcription, replication, and cellular differentiation. Upon differentiation into effector and/or memory CTLs, the majority of these gene loci lose repressive H3K27me3 while retaining the permissive H3K4me3 modification. In contrast, immune-related effector gene promoters within naive T cells lacked the permissive H3K4me3 modification, with acquisition of this modification occurring upon differentiation into effector/memory CTLs. Thus, coordinate transcriptional regulation of CTL genes with related functions is achieved via distinct epigenetic mechanisms.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Allan RS, Zueva E, Cammas F, Schreiber HA, Masson V, Belz GT, Roche D, Maison C, Quivy JP, Almouzni G, Amigorena S. An epigenetic silencing pathway controlling T helper 2 cell lineage commitment. Nature. 2012;487:249–253. - PubMed
-
- Ansel KM, Lee DU, Rao A. An epigenetic view of helper T cell differentiation. Nat Immunol. 2003;4:616–623. - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
