

Role of Cdk1 in the p53-independent abrogation of the postmitotic checkpoint by human papillomavirus E6
- PMID: 25520504
- PMCID: PMC4325723
- DOI: 10.1128/JVI.02269-14
Role of Cdk1 in the p53-independent abrogation of the postmitotic checkpoint by human papillomavirus E6
Abstract
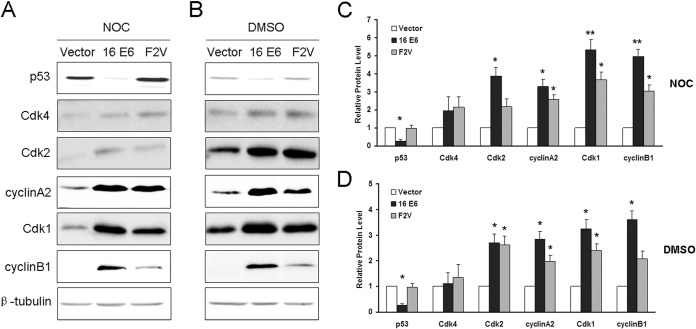
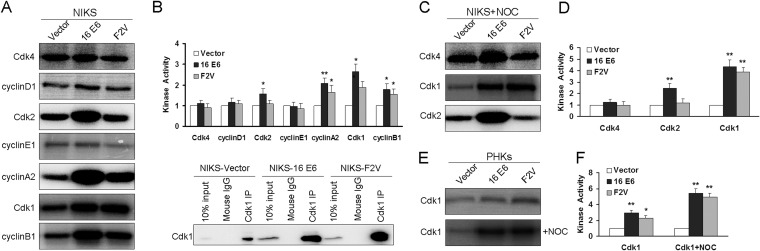
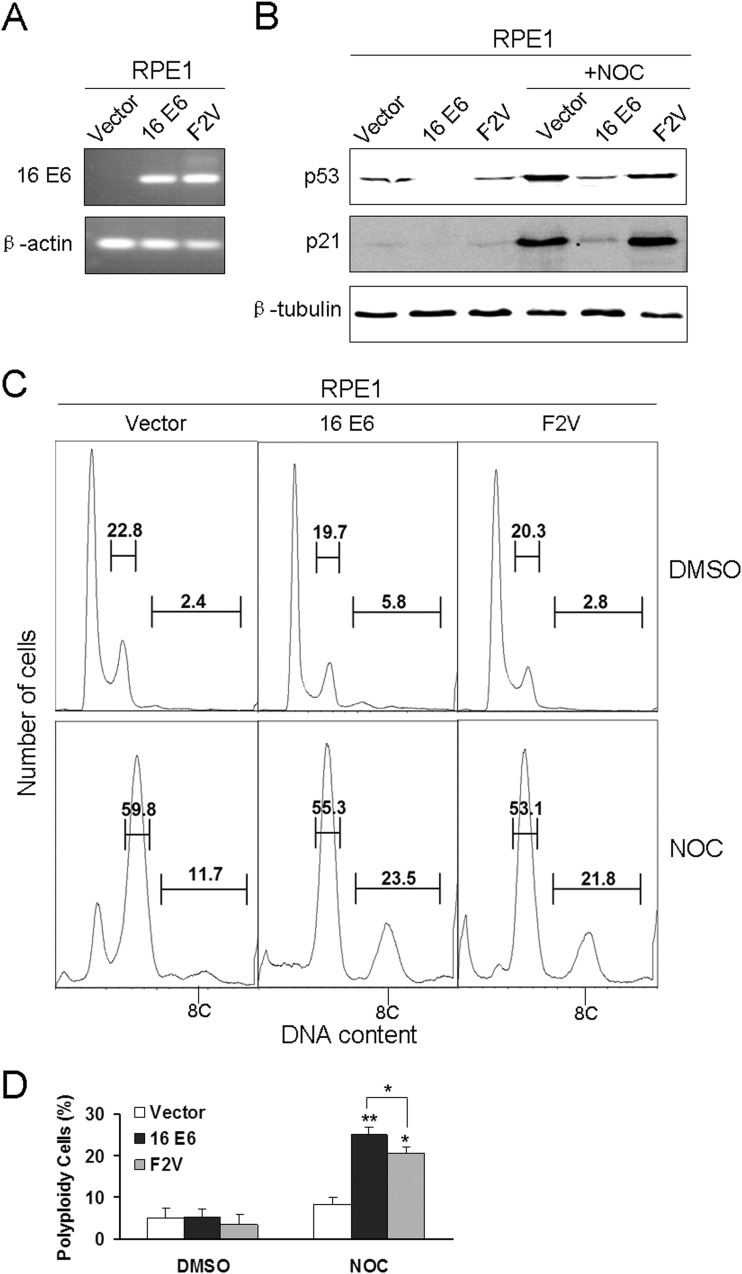
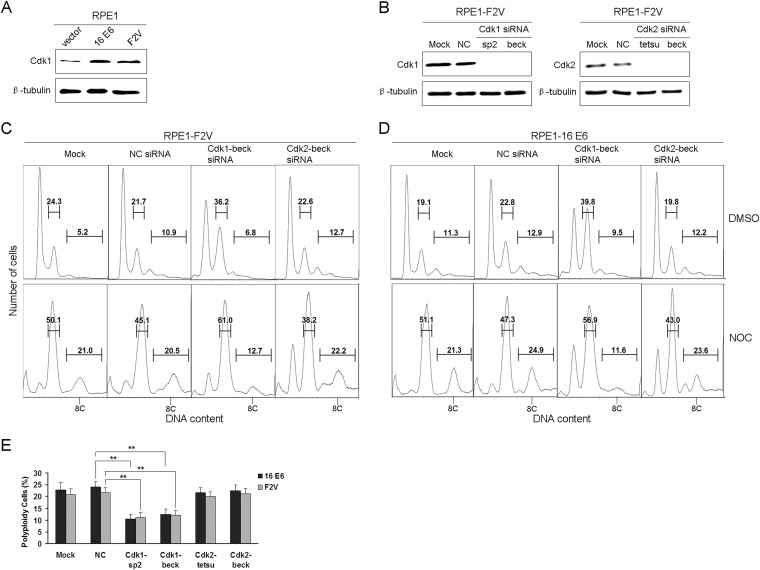
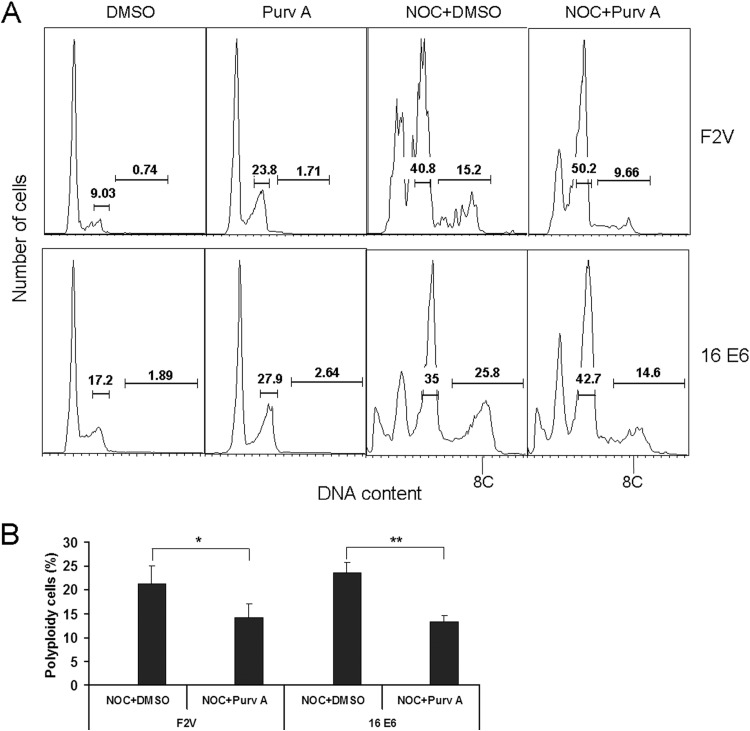
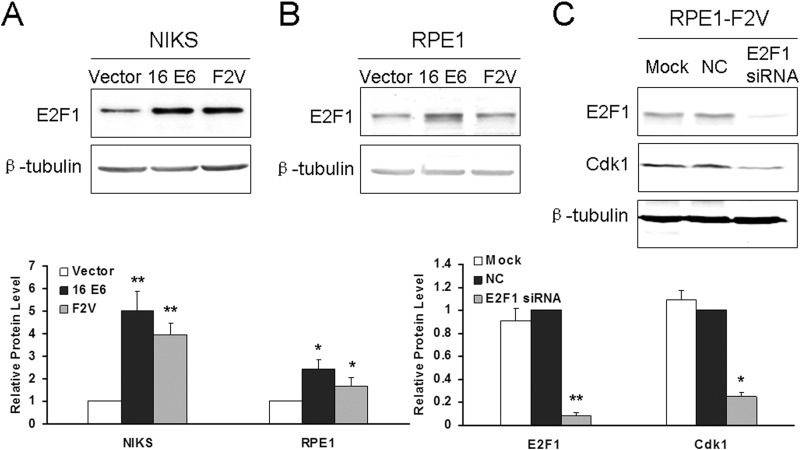
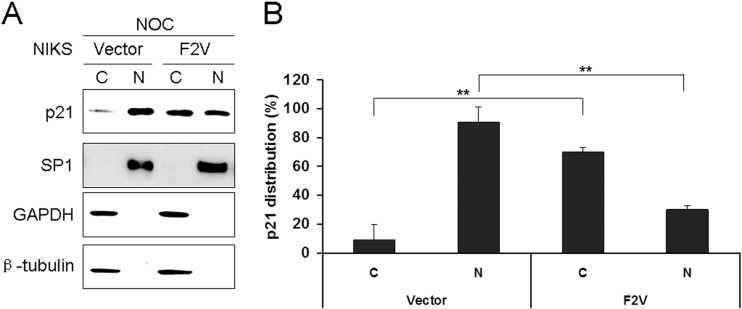
Specific types of human papillomavirus (HPV) are strongly associated with the development of cervical carcinoma. The HPV E6 oncoprotein from HPV degrades p53 and abrogates cell cycle checkpoints. Nonetheless, functional p53 has been observed in cervical cancer. We have previously identified a p53-independent function of E6 in attenuating the postmitotic G1-like checkpoint that can lead to polyploidy, an early event during cervical carcinogenesis that predisposes cells to aneuploidy. How E6 promotes cell cycle progression in the presence of p53 and its target, p21, remains a mystery. In this study, we examined the expression of cell cycle-related genes in cells expressing wild-type E6 and the mutant that is defective in p53 degradation but competent in abrogating the postmitotic checkpoint. Our results demonstrated an increase in the steady-state levels of G1- and G2-related cyclins/Cdks in E6-expressing keratinocytes. Interestingly, only Cdk1 remained active in E6 mutant-expressing cells while bypassing the postmitotic checkpoint. Furthermore, the downregulation of Cdk1 impaired the ability of both wild-type and mutant E6 to induce polyploidy. Our study thus demonstrated an important role for Cdk1, which binds p21 with lower affinity than Cdk2, in abrogating the postmitotic checkpoint in E6-expressing cells. We further show that E2F1 is important for E6 to upregulate Cdk1. Moreover, reduced nuclear p21 localization was observed in the E6 mutant-expressing cells. These findings shed light on the mechanisms by which HPV induces genomic instability and hold promise for the identification of drug targets.
Importance: HPV infection is strongly associated with the development of cervical carcinoma. HPV encodes an E6 oncoprotein that degrades the tumor suppressor p53 and abrogates cell cycle checkpoints. Nonetheless, functional p53 has been observed in cervical cancer. We have recently demonstrated a p53-independent abrogation of the postmitotic checkpoint by HPV E6 that induces polyploidy. However, the mechanism is not known. In this study, we provide evidence that Cdk1 plays an important role in this process. Previously, Cdk2 was thought to be essential for the G1/S transition, while Cdk1 only compensated its function in the absence of Cdk2. Our studies have demonstrated a novel role of Cdk1 at the postmitotic G1-like checkpoint in the presence of Cdk2. These findings shed light on the mechanisms by which HPV induces genomic instability and hold promise for the identification of drug targets.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
p53-independent abrogation of a postmitotic checkpoint contributes to human papillomavirus E6-induced polyploidy.Cancer Res. 2007 Mar 15;67(6):2603-10. doi: 10.1158/0008-5472.CAN-06-3436. Cancer Res. 2007. PMID: 17363579
-
CIP2A facilitates the G1/S cell cycle transition via B-Myb in human papillomavirus 16 oncoprotein E6-expressing cells.J Cell Mol Med. 2018 Sep;22(9):4150-4160. doi: 10.1111/jcmm.13693. Epub 2018 Jun 12. J Cell Mol Med. 2018. PMID: 29893470 Free PMC article.
-
Role of Cdk1 in DNA damage-induced G1 checkpoint abrogation by the human papillomavirus E7 oncogene.Cell Cycle. 2014;13(20):3249-59. doi: 10.4161/15384101.2014.953879. Cell Cycle. 2014. PMID: 25485505 Free PMC article.
-
The role of TP53 in Cervical carcinogenesis.Hum Mutat. 2003 Mar;21(3):307-12. doi: 10.1002/humu.10178. Hum Mutat. 2003. PMID: 12619117 Review.
-
Regulation of cell cycle progression and apoptosis by the papillomavirus E6 oncogene.Crit Rev Eukaryot Gene Expr. 2004;14(3):183-202. doi: 10.1615/critreveukaryotgeneexpr.v14.i3.30. Crit Rev Eukaryot Gene Expr. 2004. PMID: 15248815 Review.
Cited by
-
Bioinformatics prediction and analysis of hub genes and pathways of three types of gynecological cancer.Oncol Lett. 2019 Jul;18(1):617-628. doi: 10.3892/ol.2019.10371. Epub 2019 May 17. Oncol Lett. 2019. PMID: 31289534 Free PMC article.
-
Effects of a Closed Space Environment on Gene Expression in Hair Follicles of Astronauts in the International Space Station.PLoS One. 2016 Mar 30;11(3):e0150801. doi: 10.1371/journal.pone.0150801. eCollection 2016. PLoS One. 2016. PMID: 27029003 Free PMC article.
-
Polyploid Giant Cancer Cells, a Hallmark of Oncoviruses and a New Therapeutic Challenge.Front Oncol. 2020 Oct 14;10:567116. doi: 10.3389/fonc.2020.567116. eCollection 2020. Front Oncol. 2020. PMID: 33154944 Free PMC article. Review.
-
Systematic assessment of cervical cancer initiation and progression uncovers genetic panels for deep learning-based early diagnosis and proposes novel diagnostic and prognostic biomarkers.Oncotarget. 2017 Nov 25;8(65):109436-109456. doi: 10.18632/oncotarget.22689. eCollection 2017 Dec 12. Oncotarget. 2017. PMID: 29312619 Free PMC article.
-
Synergistic cytotoxic effects of a combined treatment of a Pinellia pedatisecta lipid-soluble extract and cisplatin on human cervical carcinoma in vivo.Oncol Lett. 2017 Jun;13(6):4748-4754. doi: 10.3892/ol.2017.6091. Epub 2017 Apr 25. Oncol Lett. 2017. PMID: 28588727 Free PMC article.
References
-
- Giannoudis A, Evans MF, Southern SA, Herrington CS. 2000. Basal keratinocyte tetrasomy in low-grade squamous intra-epithelial lesions of the cervix is restricted to high and intermediate risk HPV infection but is not type-specific. Brit J Cancer 82:424–428. doi:10.1054/bjoc.1999.0937. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
