

"You Shall Not Pass"-tight junctions of the blood brain barrier
- PMID: 25520612
- PMCID: PMC4253952
- DOI: 10.3389/fnins.2014.00392
"You Shall Not Pass"-tight junctions of the blood brain barrier
Abstract
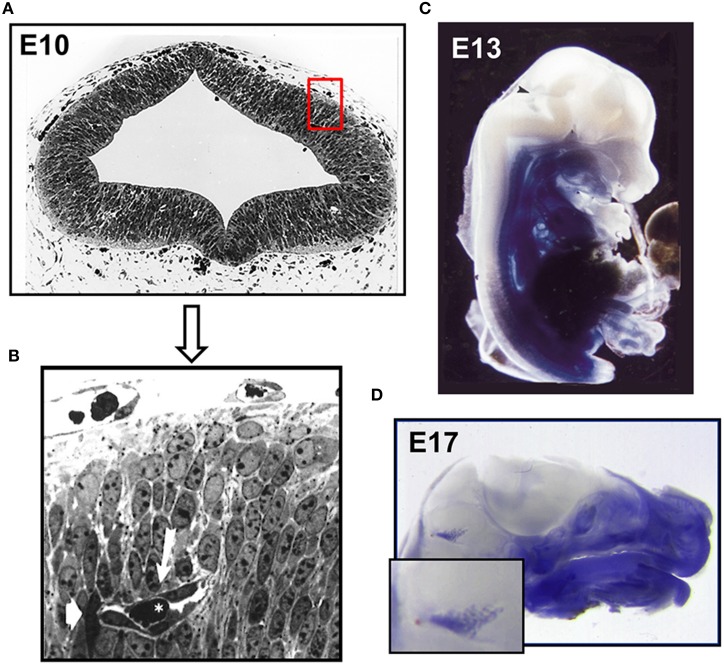
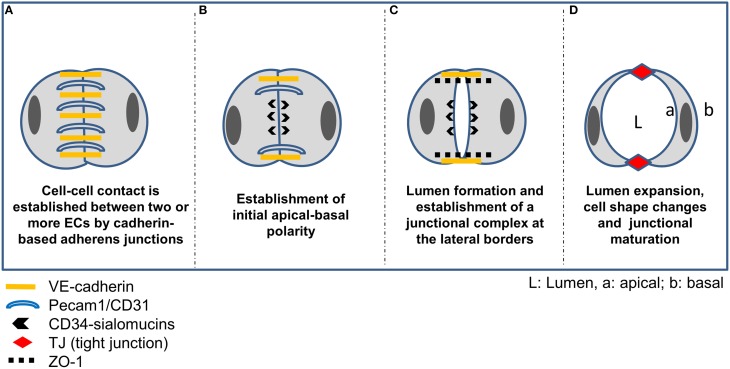
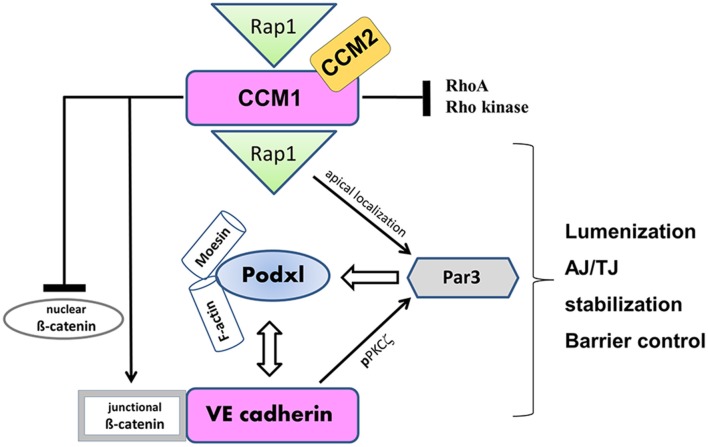
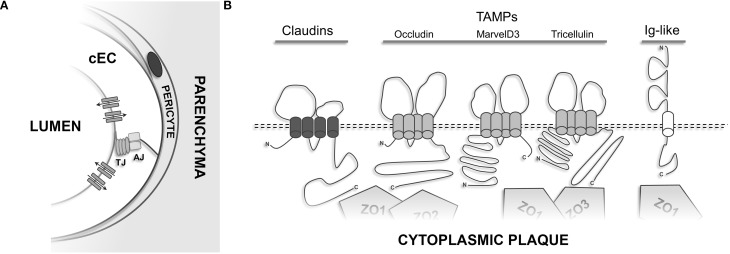
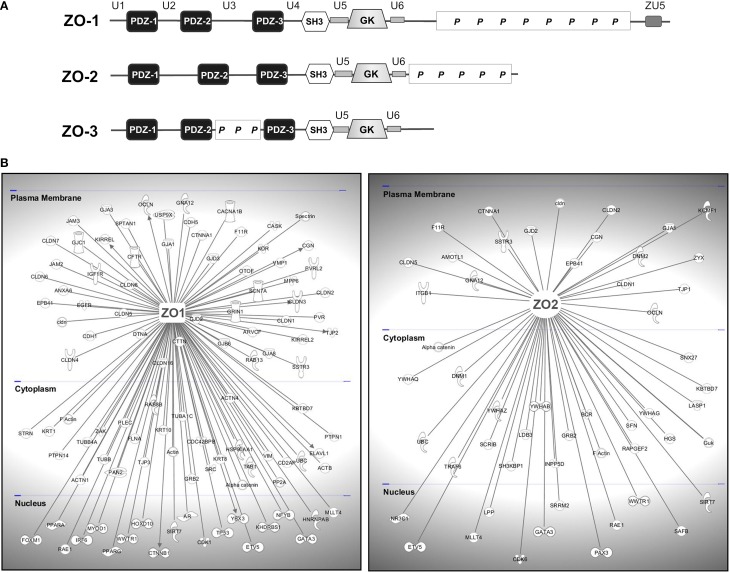
The structure and function of the barrier layers restricting the free diffusion of substances between the central nervous system (brain and spinal cord) and the systemic circulation is of great medical interest as various pathological conditions often lead to their impairment. Excessive leakage of blood-borne molecules into the parenchyma and the concomitant fluctuations in the microenvironment following a transient breakdown of the blood-brain barrier (BBB) during ischemic/hypoxic conditions or because of an autoimmune disease are detrimental to the physiological functioning of nervous tissue. On the other hand, the treatment of neurological disorders is often hampered as only minimal amounts of therapeutic agents are able to penetrate a fully functional BBB or blood cerebrospinal fluid barrier. An in-depth understanding of the molecular machinery governing the establishment and maintenance of these barriers is necessary to develop rational strategies allowing a controlled delivery of appropriate drugs to the CNS. At the basis of such tissue barriers are intimate cell-cell contacts (zonulae occludentes, tight junctions) which are present in all polarized epithelia and endothelia. By creating a paracellular diffusion constraint TJs enable the vectorial transport across cell monolayers. More recent findings indicate that functional barriers are already established during development, protecting the fetal brain. As an understanding of the biogenesis of TJs might reveal the underlying mechanisms of barrier formation during ontogenic development numerous in vitro systems have been developed to study the assembly and disassembly of TJs. In addition, monitoring the stage-specific expression of TJ-associated proteins during development has brought much insight into the "developmental tightening" of tissue barriers. Over the last two decades a detailed molecular map of transmembrane and cytoplasmic TJ-proteins has been identified. These proteins not only form a cell-cell adhesion structure, but integrate various signaling pathways, thereby directly or indirectly impacting upon processes such as cell-cell adhesion, cytoskeletal rearrangement, and transcriptional control. This review will provide a brief overview on the establishment of the BBB during embryonic development in mammals and a detailed description of the ultrastructure, biogenesis, and molecular composition of epithelial and endothelial TJs will be given.
Keywords: MAGUK proteins; PDZ scaffolds; blood-brain barrier; brain capillary endothelial cells; cell polarity; tight junctions; vascular permeability.
Figures
References
-
- Adamson P., Etienne S., Couraud P. O., Calder V., Greenwood J. (1999). Lymphocyte migration through brain endothelial cell monolayers involves signaling through endothelial ICAM-1 via a rho-dependent pathway. J. Immunol. 162, 2964–2973. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials