

Mutations in KATNB1 cause complex cerebral malformations by disrupting asymmetrically dividing neural progenitors
- PMID: 25521378
- PMCID: PMC5024344
- DOI: 10.1016/j.neuron.2014.12.014
Mutations in KATNB1 cause complex cerebral malformations by disrupting asymmetrically dividing neural progenitors
Erratum in
- Neuron. 2015 Jan 7;85(1):228
-
Mutations in KATNB1 Cause Complex Cerebral Malformations by Disrupting Asymmetrically Dividing Neural Progenitors.Neuron. 2015 Jan 7;85(1):228. doi: 10.1016/j.neuron.2014.12.046. Epub 2015 Jan 7. Neuron. 2015. PMID: 29654772 No abstract available.
Abstract
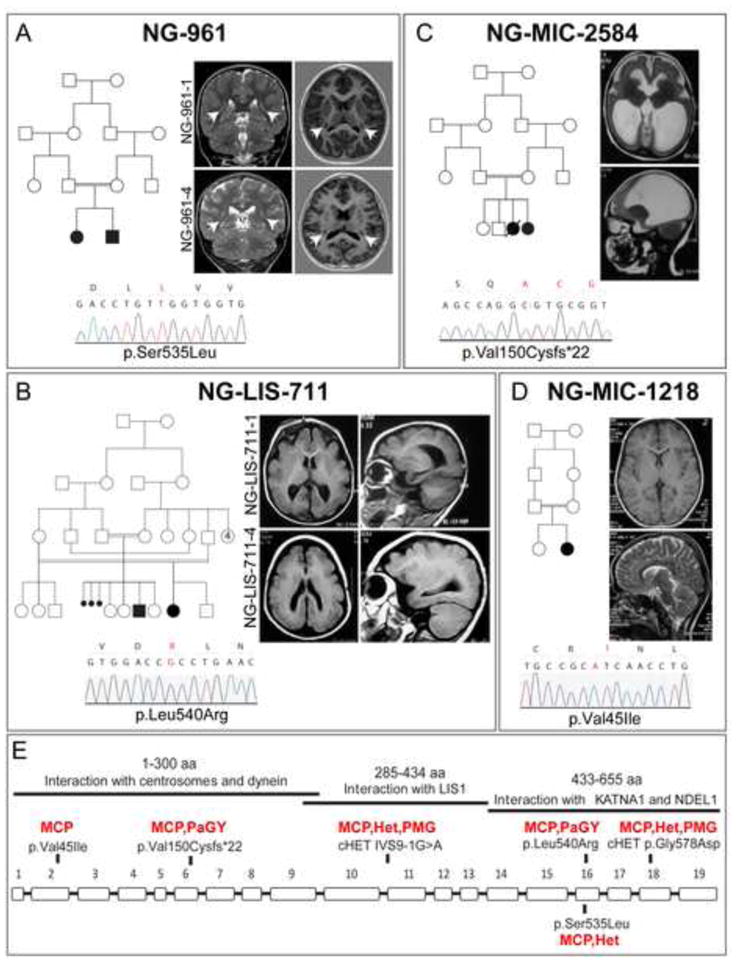
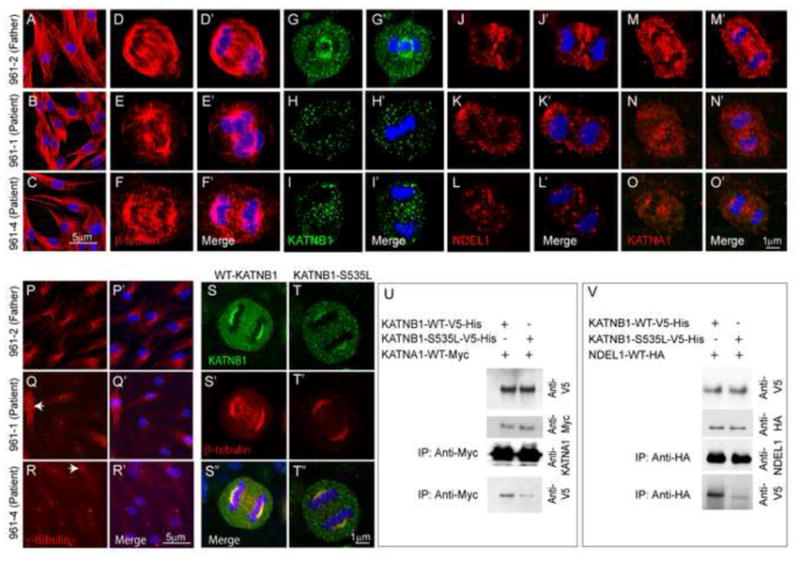
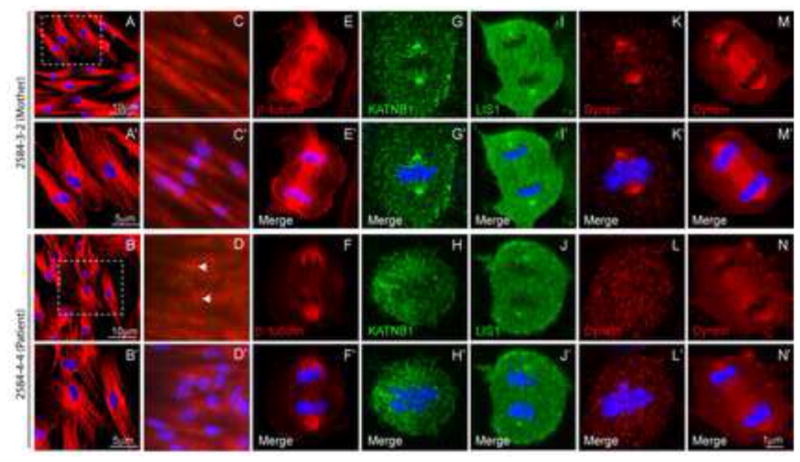
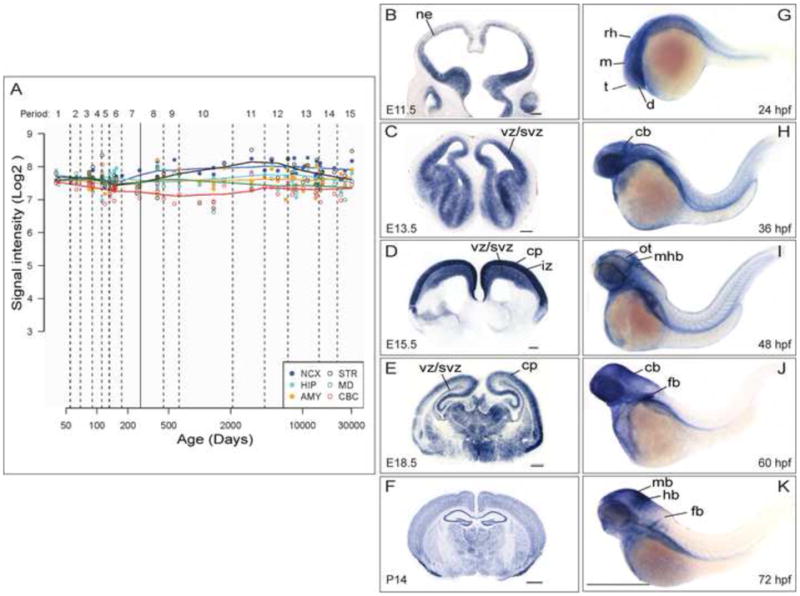
Exome sequencing analysis of over 2,000 children with complex malformations of cortical development identified five independent (four homozygous and one compound heterozygous) deleterious mutations in KATNB1, encoding the regulatory subunit of the microtubule-severing enzyme Katanin. Mitotic spindle formation is defective in patient-derived fibroblasts, a consequence of disrupted interactions of mutant KATNB1 with KATNA1, the catalytic subunit of Katanin, and other microtubule-associated proteins. Loss of KATNB1 orthologs in zebrafish (katnb1) and flies (kat80) results in microcephaly, recapitulating the human phenotype. In the developing Drosophila optic lobe, kat80 loss specifically affects the asymmetrically dividing neuroblasts, which display supernumerary centrosomes and spindle abnormalities during mitosis, leading to cell cycle progression delays and reduced cell numbers. Furthermore, kat80 depletion results in dendritic arborization defects in sensory and motor neurons, affecting neural architecture. Taken together, we provide insight into the mechanisms by which KATNB1 mutations cause human cerebral cortical malformations, demonstrating its fundamental role during brain development.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
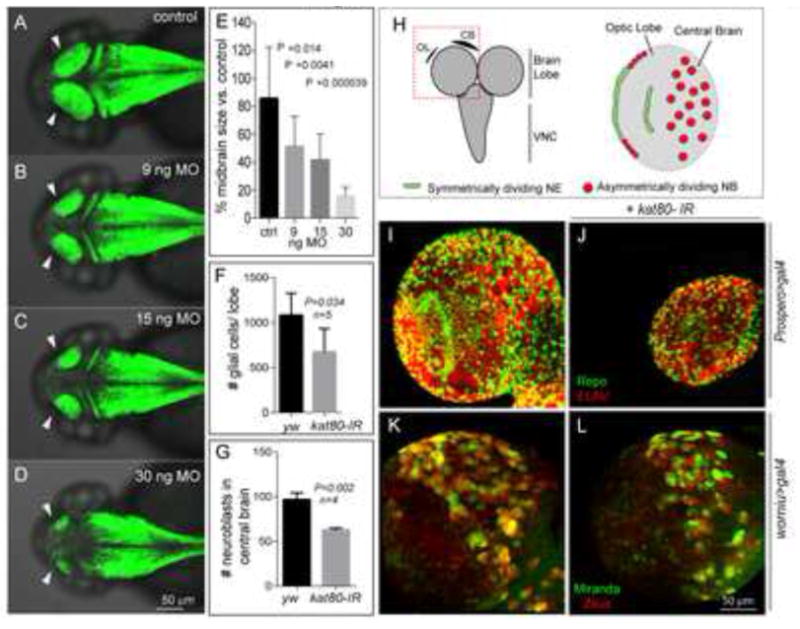
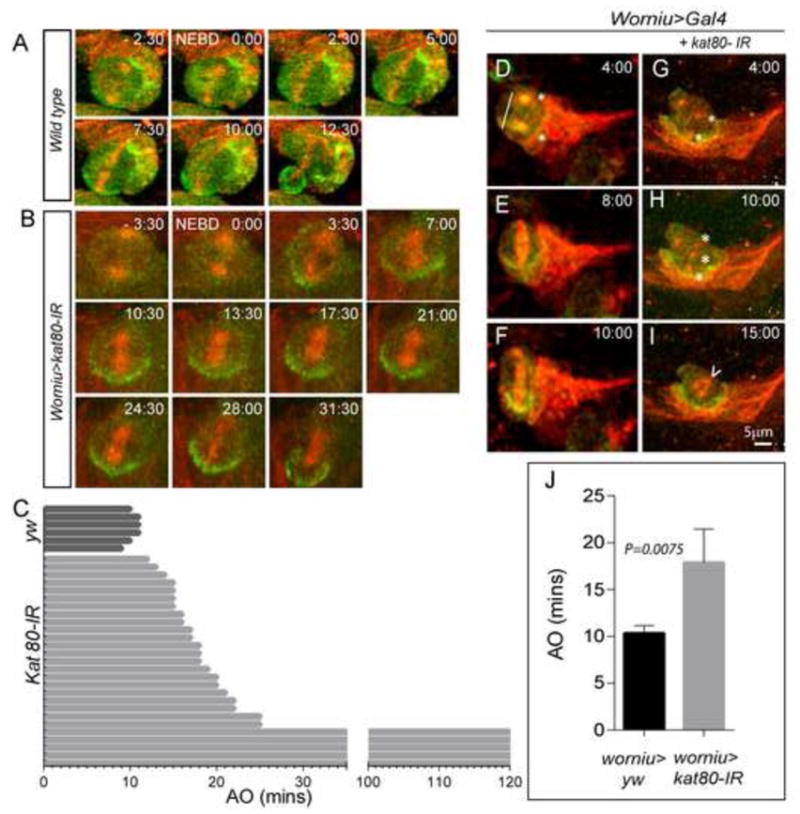
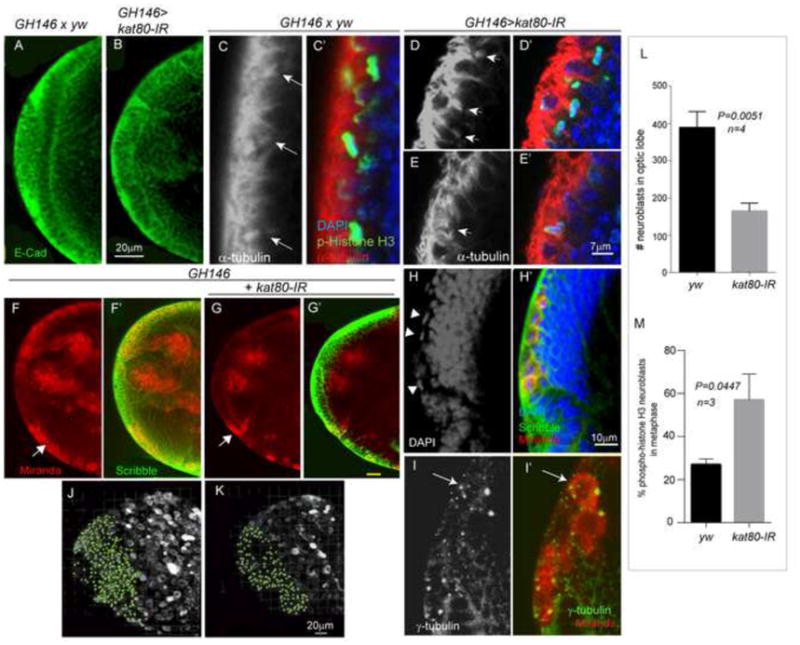
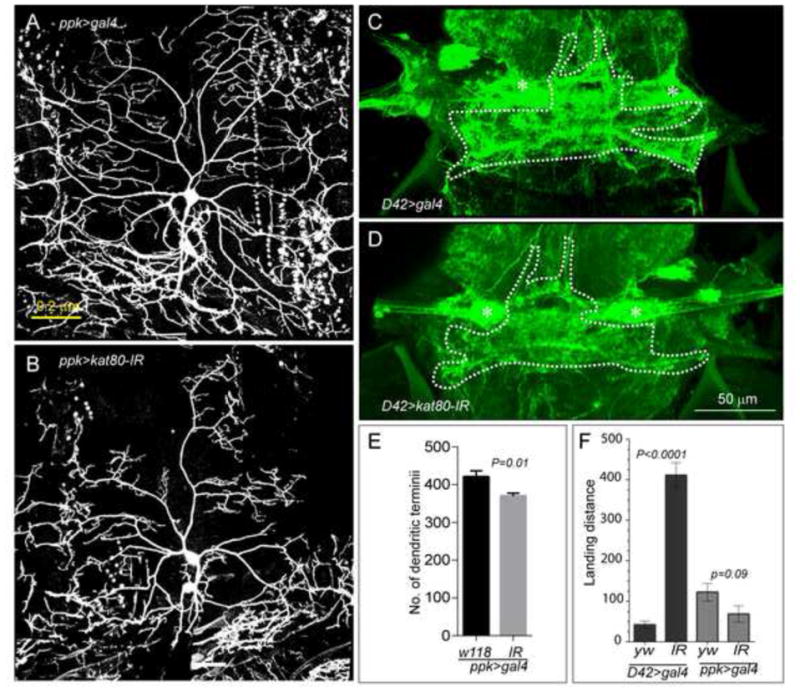
References
-
- Armstrong DD. Neuropathology of Rett syndrome. Journal of child neurology. 2005;20:747–753. - PubMed
-
- Ashraf SI, Ganguly A, Roote J, Ip YT. Worniu, a Snail family zinc-finger protein, is required for brain development in Drosophila. Developmental dynamics: an official publication of the American Association of Anatomists. 2004;231:379–386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01MH081896/MH/NIMH NIH HHS/United States
- UM1 HG006504/HG/NHGRI NIH HHS/United States
- R01 NS041537/NS/NINDS NIH HHS/United States
- R01MH103616/MH/NIMH NIH HHS/United States
- U01 MH081896/MH/NIMH NIH HHS/United States
- P01 HD070494/HD/NICHD NIH HHS/United States
- R00 HD082337/HD/NICHD NIH HHS/United States
- R01NS041537/NS/NINDS NIH HHS/United States
- U54HG006504/HG/NHGRI NIH HHS/United States
- U54 HG006504/HG/NHGRI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 CA069408/CA/NCI NIH HHS/United States
- K99 HD082337/HD/NICHD NIH HHS/United States
- P01HD070494/HD/NICHD NIH HHS/United States
- R01 MH103616/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
