

SCL, LMO1 and Notch1 reprogram thymocytes into self-renewing cells
- PMID: 25522233
- PMCID: PMC4270438
- DOI: 10.1371/journal.pgen.1004768
SCL, LMO1 and Notch1 reprogram thymocytes into self-renewing cells
Abstract
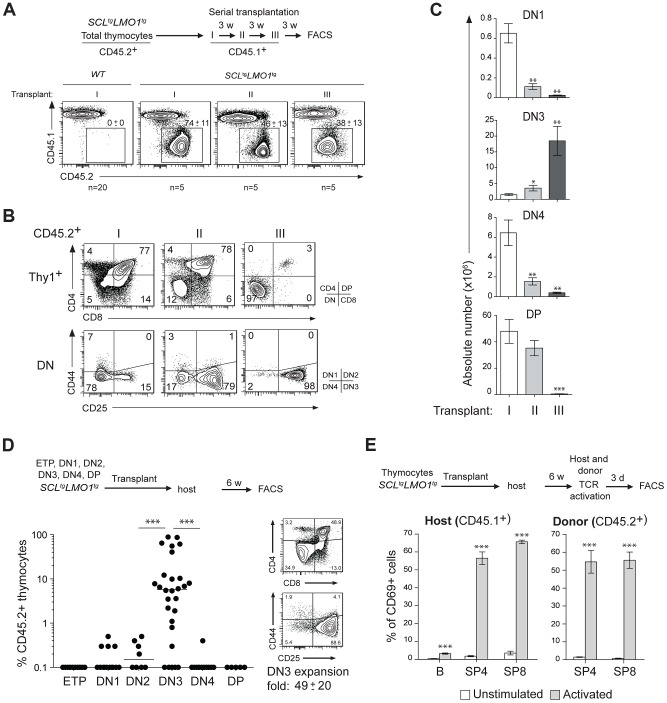
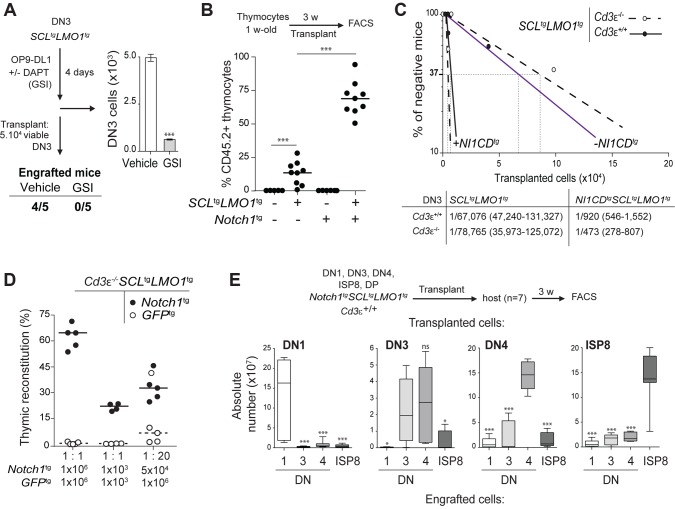
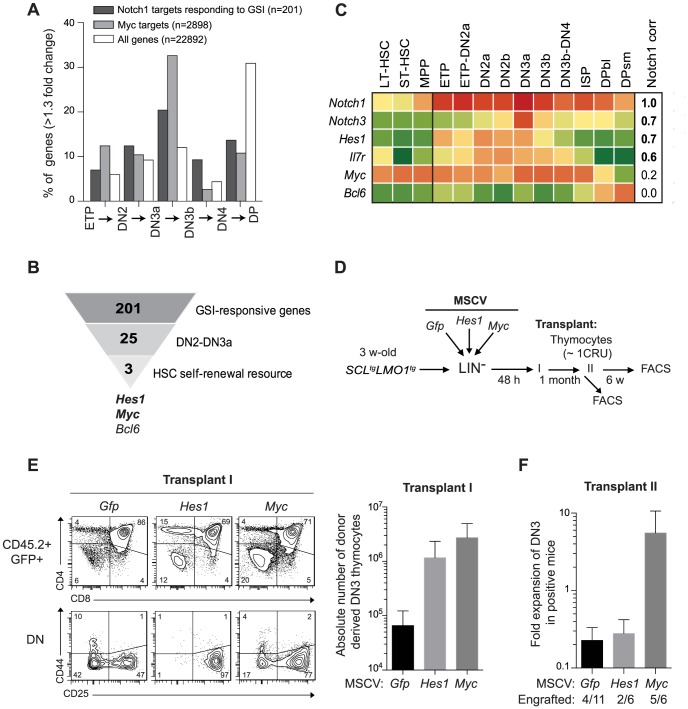
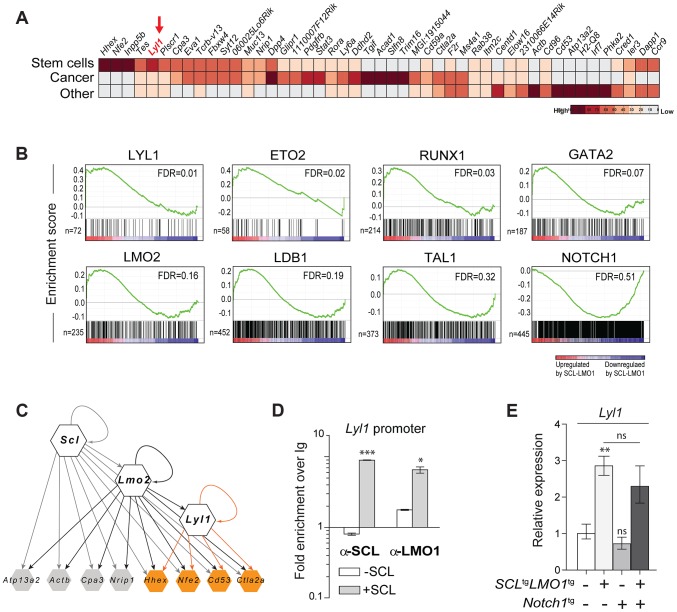
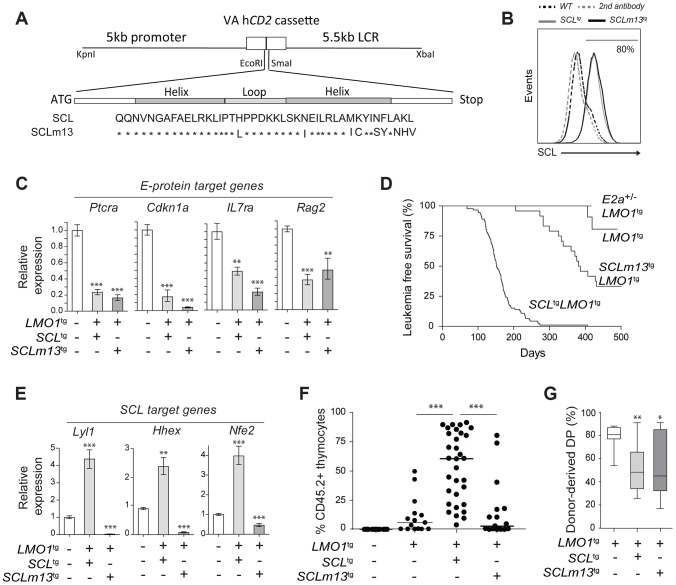
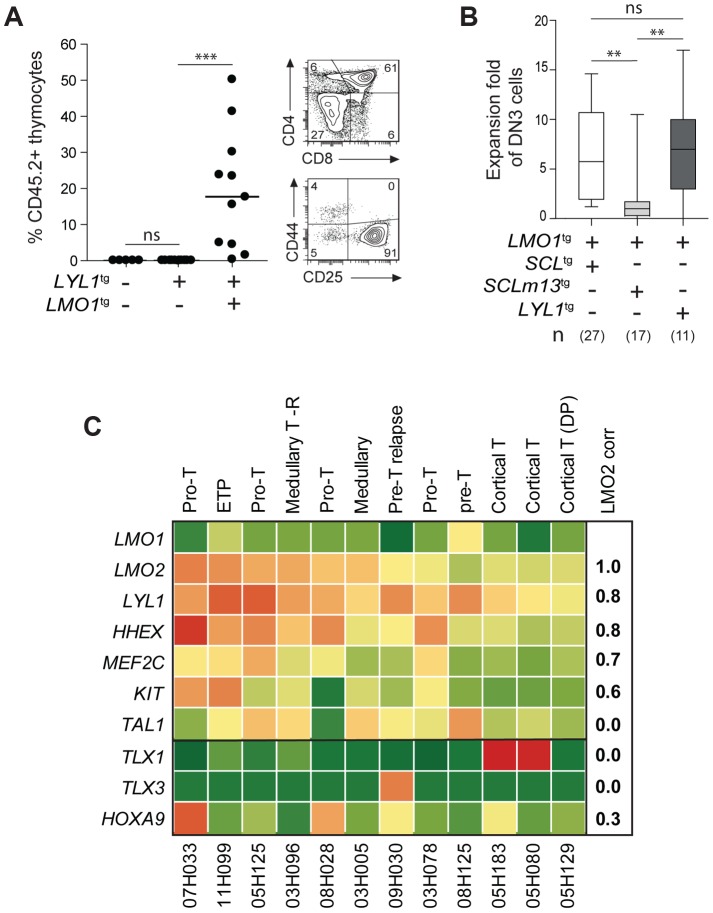
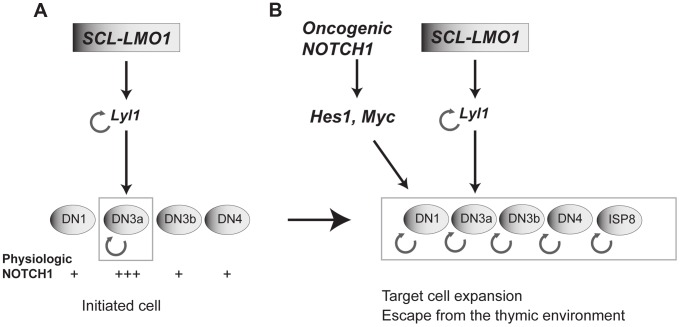
The molecular determinants that render specific populations of normal cells susceptible to oncogenic reprogramming into self-renewing cancer stem cells are poorly understood. Here, we exploit T-cell acute lymphoblastic leukemia (T-ALL) as a model to define the critical initiating events in this disease. First, thymocytes that are reprogrammed by the SCL and LMO1 oncogenic transcription factors into self-renewing pre-leukemic stem cells (pre-LSCs) remain non-malignant, as evidenced by their capacities to generate functional T cells. Second, we provide strong genetic evidence that SCL directly interacts with LMO1 to activate the transcription of a self-renewal program coordinated by LYL1. Moreover, LYL1 can substitute for SCL to reprogram thymocytes in concert with LMO1. In contrast, inhibition of E2A was not sufficient to substitute for SCL, indicating that thymocyte reprogramming requires transcription activation by SCL-LMO1. Third, only a specific subset of normal thymic cells, known as DN3 thymocytes, is susceptible to reprogramming. This is because physiological NOTCH1 signals are highest in DN3 cells compared to other thymocyte subsets. Consistent with this, overexpression of a ligand-independent hyperactive NOTCH1 allele in all immature thymocytes is sufficient to sensitize them to SCL-LMO1, thereby increasing the pool of self-renewing cells. Surprisingly, hyperactive NOTCH1 cannot reprogram thymocytes on its own, despite the fact that NOTCH1 is activated by gain of function mutations in more than 55% of T-ALL cases. Rather, elevating NOTCH1 triggers a parallel pathway involving Hes1 and Myc that dramatically enhances the activity of SCL-LMO1 We conclude that the acquisition of self-renewal and the genesis of pre-LSCs from thymocytes with a finite lifespan represent a critical first event in T-ALL. Finally, LYL1 and LMO1 or LMO2 are co-expressed in most human T-ALL samples, except the cortical T subtype. We therefore anticipate that the self-renewal network described here may be relevant to a majority of human T-ALL.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, et al. (1994) A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 367: 645–648. - PubMed
-
- Valent P, Bonnet D, Wohrer S, Andreeff M, Copland M, et al. (2013) Heterogeneity of neoplastic stem cells: theoretical, functional, and clinical implications. Cancer Res 73: 1037–1045. - PubMed
-
- Bonnet D, Dick JE (1997) Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med 3: 730–737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
