

Once upon a time the cell membranes: 175 years of cell boundary research
- PMID: 25522740
- PMCID: PMC4304622
- DOI: 10.1186/s13062-014-0032-7
Once upon a time the cell membranes: 175 years of cell boundary research
Abstract
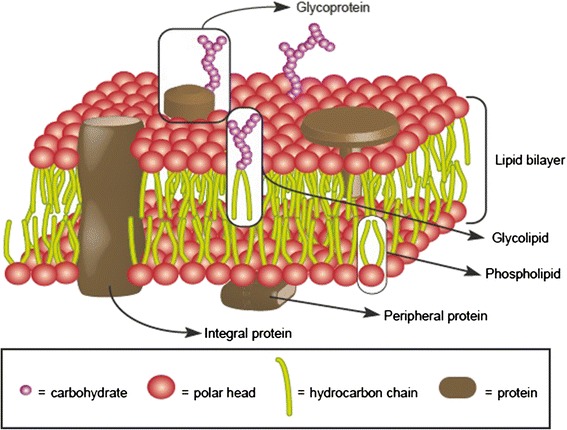
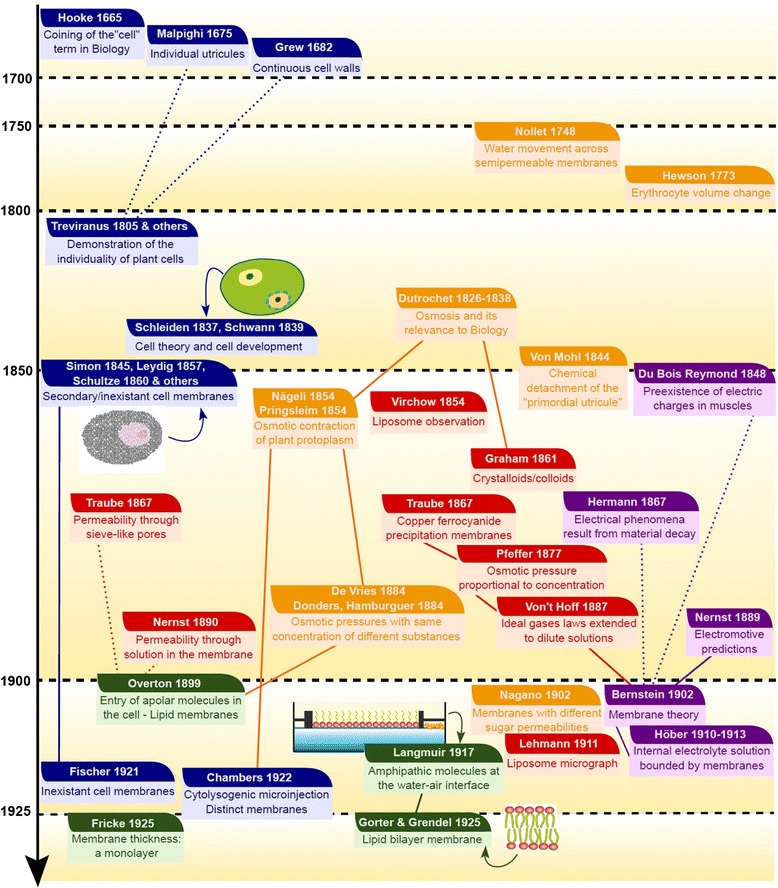
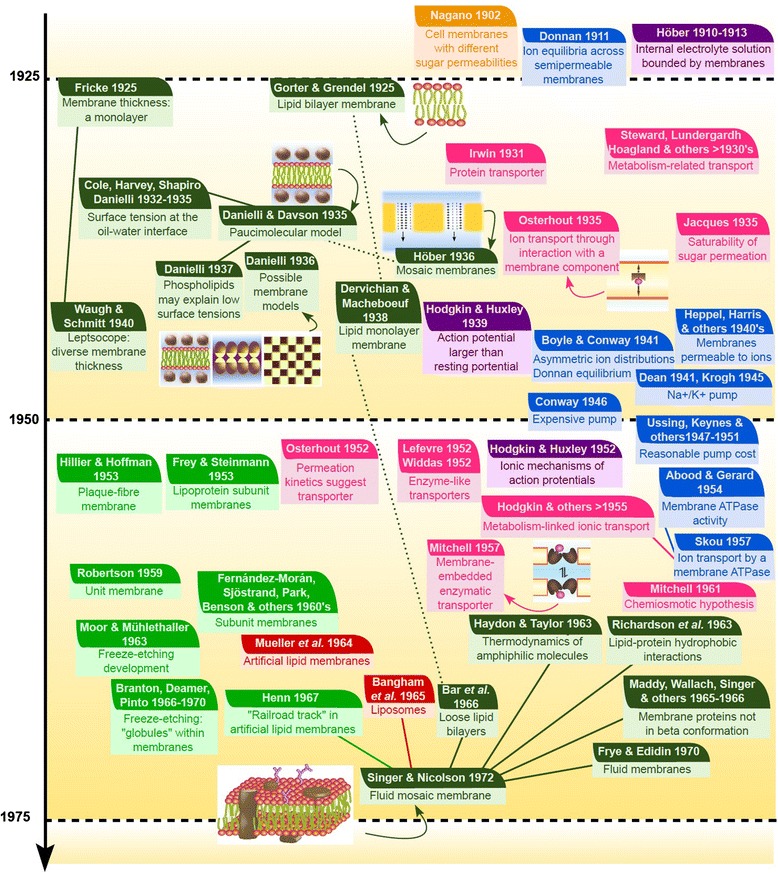
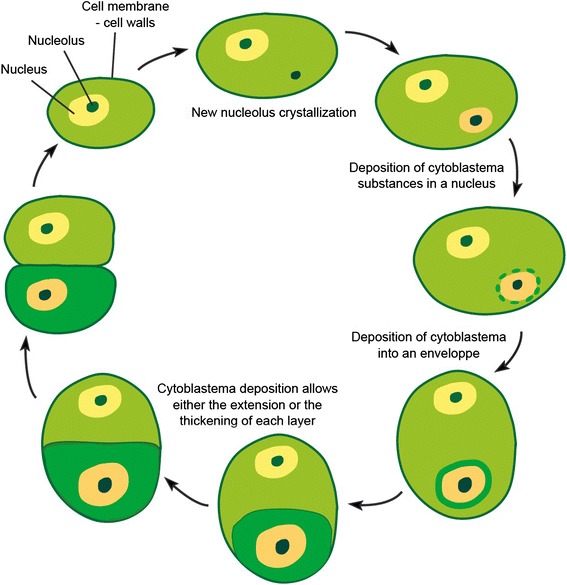
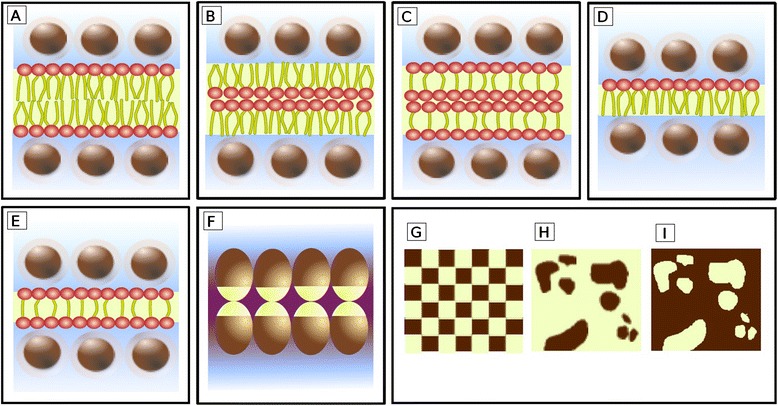
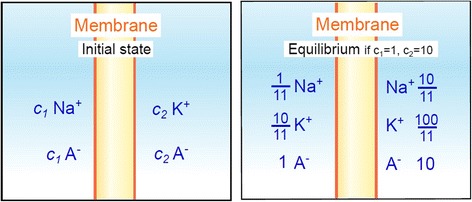
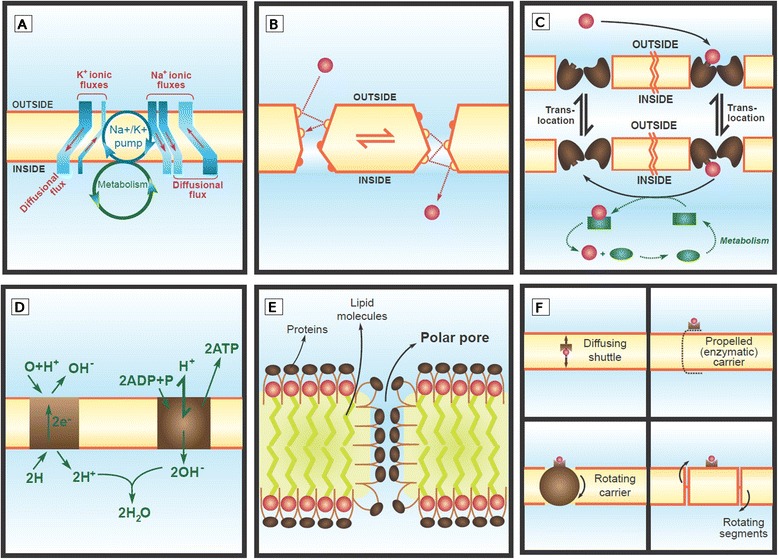
All modern cells are bounded by cell membranes best described by the fluid mosaic model. This statement is so widely accepted by biologists that little attention is generally given to the theoretical importance of cell membranes in describing the cell. This has not always been the case. When the Cell Theory was first formulated in the XIX(th) century, almost nothing was known about the cell membranes. It was not until well into the XX(th) century that the existence of the plasma membrane was broadly accepted and, even then, the fluid mosaic model did not prevail until the 1970s. How were the cell boundaries considered between the articulation of the Cell Theory around 1839 and the formulation of the fluid mosaic model that has described the cell membranes since 1972? In this review I will summarize the major historical discoveries and theories that tackled the existence and structure of membranes and I will analyze how these theories impacted the understanding of the cell. Apart from its purely historical relevance, this account can provide a starting point for considering the theoretical significance of membranes to the definition of the cell and could have implications for research on early life.
Figures
References
-
- Tanford C. The Hydrophobic Effect. Formation of Micelles and Biological Membranes. New York: Wiley; 1973.
-
- Daleke DL, Lyles JV. Identification and purification of aminophopholipid flippases. Biochem Biophys Acta. 2000;1486:108–127. - PubMed
-
- Ling GN. Debunking the alleged resurrection of the sodium pump hypothesis. Physiol Chem Phys Med NMR. 1997;29:123–198. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
