

Perinatal vs genetic programming of serotonin states associated with anxiety
- PMID: 25523893
- PMCID: PMC4397404
- DOI: 10.1038/npp.2014.331
Perinatal vs genetic programming of serotonin states associated with anxiety
Abstract
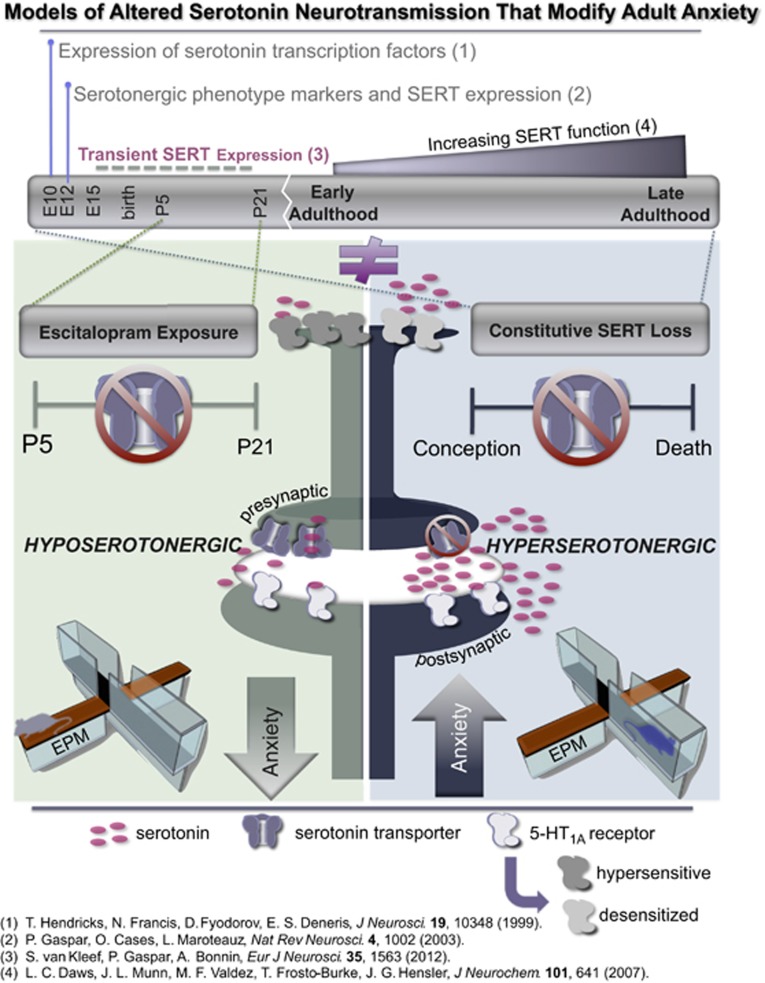
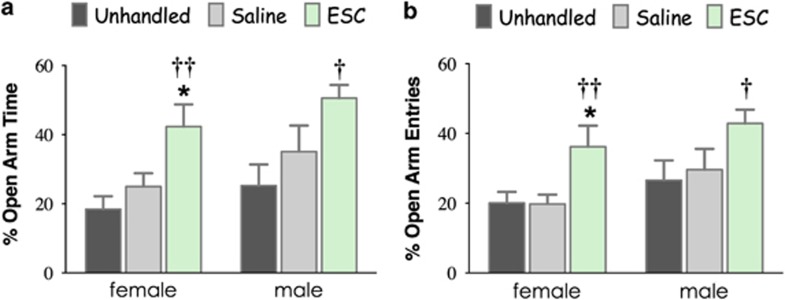
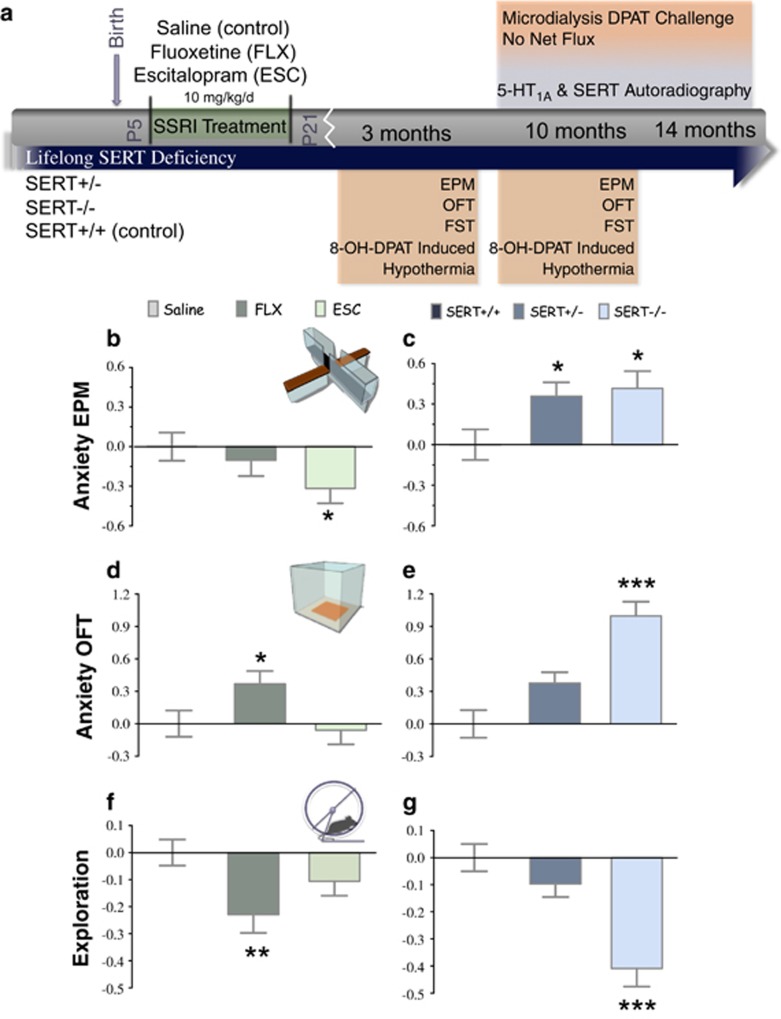
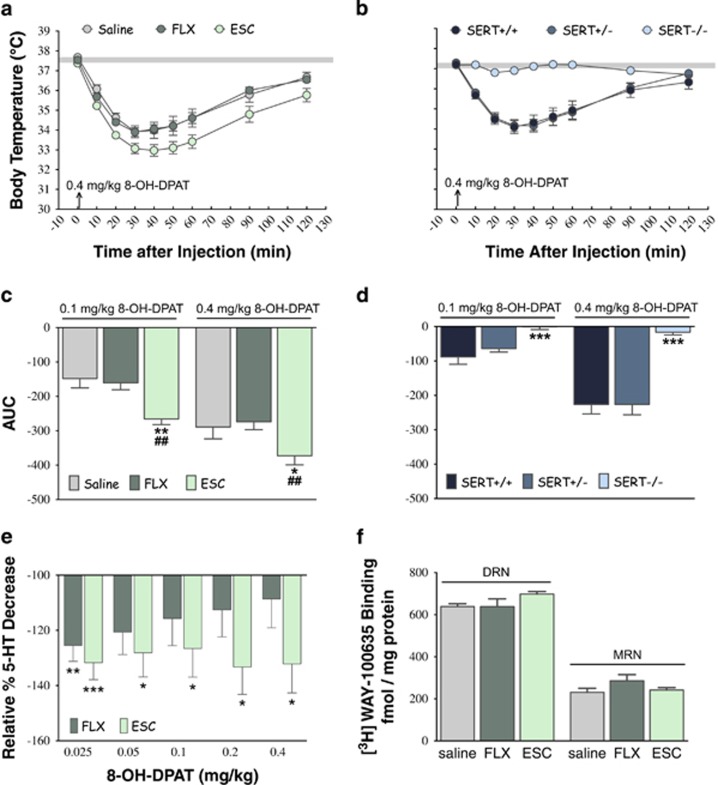
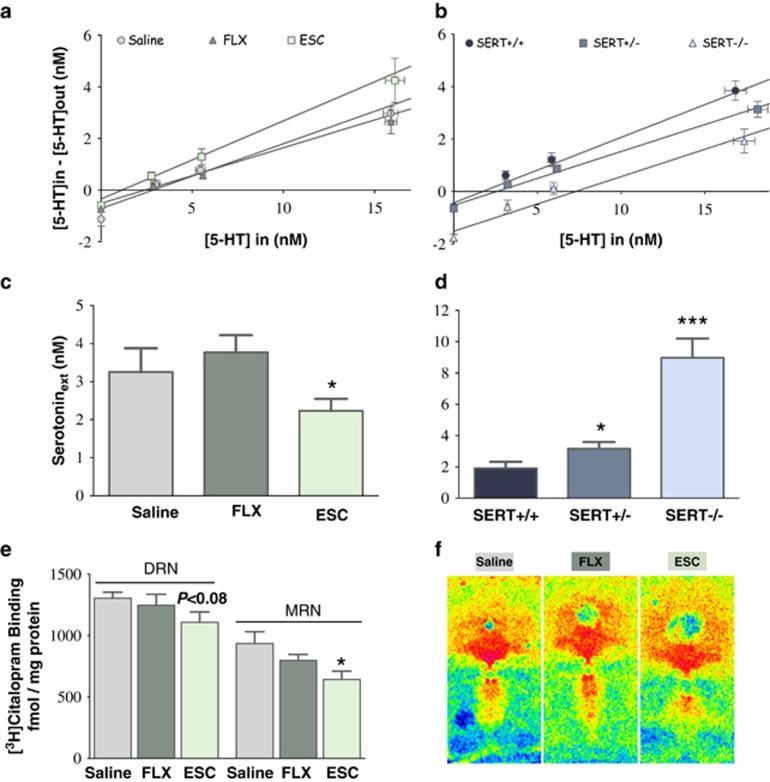
Large numbers of women undergo antidepressant treatment during pregnancy; however, long-term consequences for their offspring remain largely unknown. Rodents exposed to serotonin transporter (SERT)-inhibiting antidepressants during development show changes in adult emotion-like behavior. These changes have been equated with behavioral alterations arising from genetic reductions in SERT. Both models are highly relevant to humans yet they vary in their time frames of SERT disruption. We find that anxiety-related behavior and, importantly, underlying serotonin neurotransmission diverge between the two models. In mice, constitutive loss of SERT causes life-long increases in anxiety-related behavior and hyperserotonemia. Conversely, early exposure to the antidepressant escitalopram (ESC; Lexapro) results in decreased anxiety-related behavior beginning in adolescence, which is associated with adult serotonin system hypofunction in the ventral hippocampus. Adult behavioral changes resulting from early fluoxetine (Prozac) exposure were different from those of ESC and, although somewhat similar to SERT deficiency, were not associated with changes in hippocampal serotonin transmission in late adulthood. These findings reveal dissimilarities in adult behavior and neurotransmission arising from developmental exposure to different widely prescribed antidepressants that are not recapitulated by genetic SERT insufficiency. Moreover, they support a pivotal role for serotonergic modulation of anxiety-related behavior.
Figures
References
-
- Altieri S, Singh Y, Sibille E, Andrews A.2012Serotonergic pathways in depressionIn: Lopez-Munoz F, Alamo C (eds). Neurobiology of Depression CRC Press, Taylor & Francis Group: Boca Raton, FL, USA; 143–170.
-
- Ansorge MS, Zhou M, Lira A, Hen R, Gingrich JA. Early-life blockade of the 5-HT transporter alters emotional behavior in adult mice. Science. 2004;306:879–881. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
