

The structural basis for specificity in lipoxygenase catalysis
- PMID: 25524168
- PMCID: PMC4353356
- DOI: 10.1002/pro.2626
The structural basis for specificity in lipoxygenase catalysis
Abstract
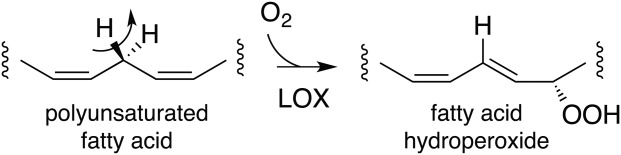
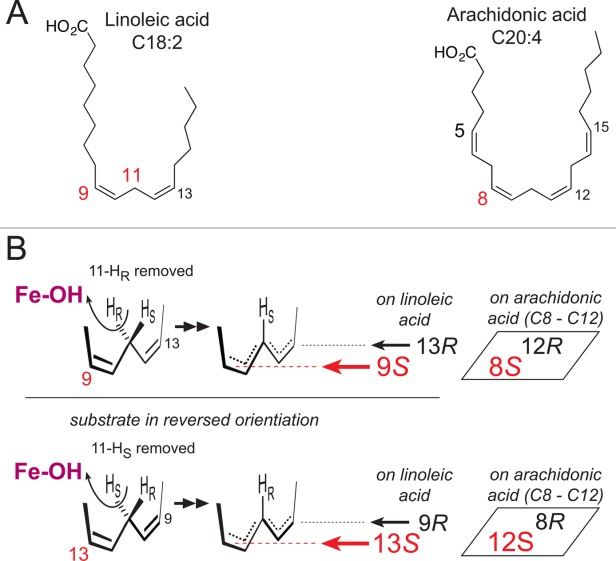
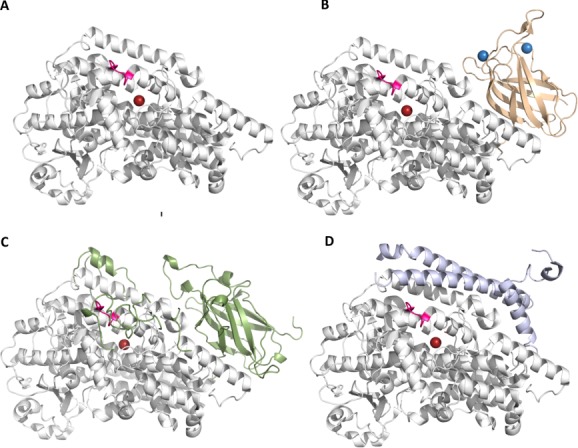
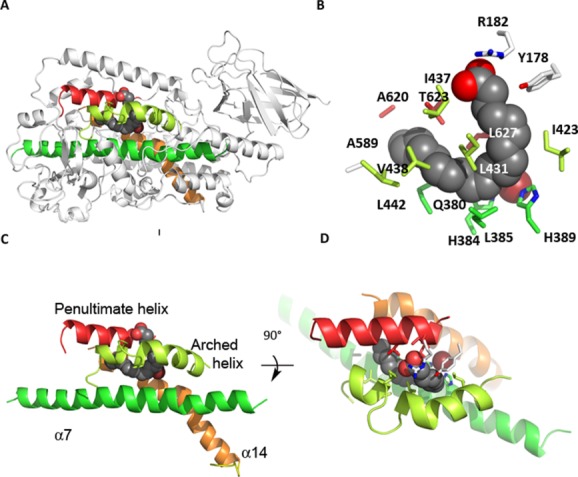
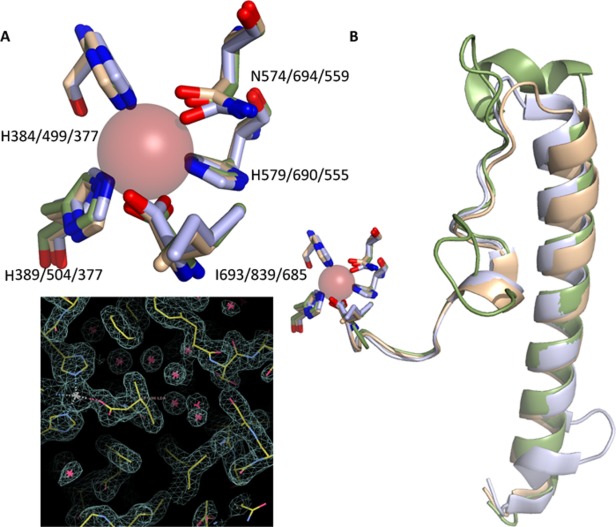
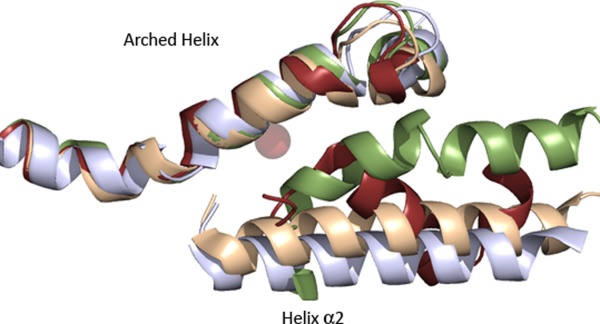
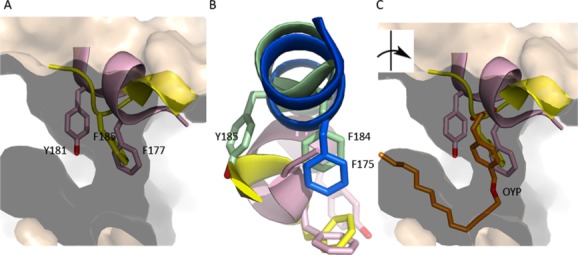
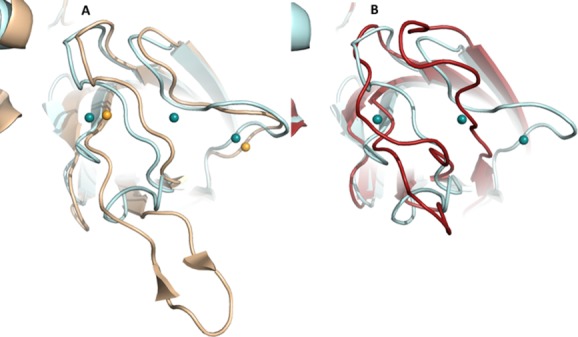
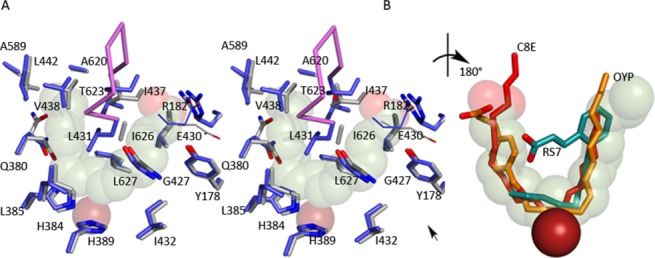
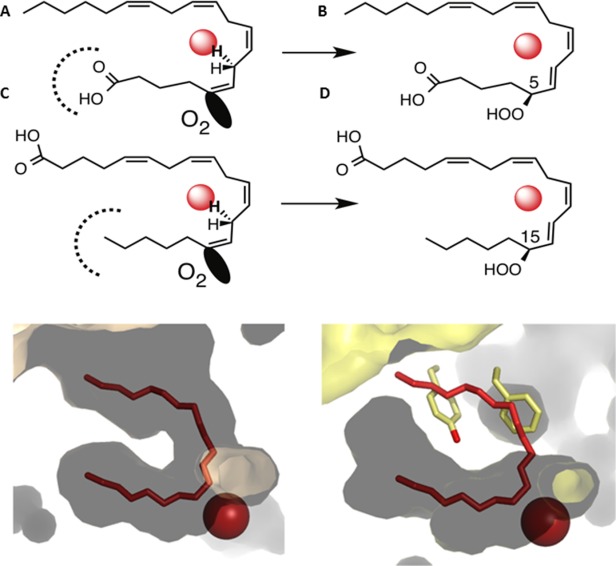
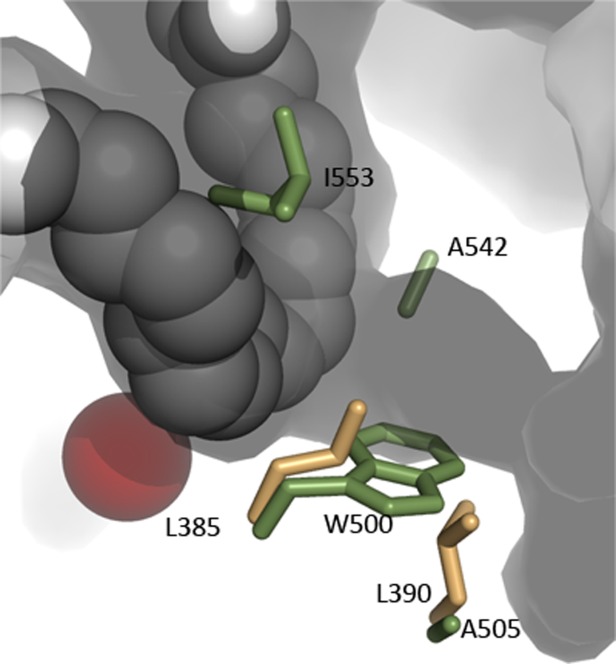
Many intriguing facets of lipoxygenase (LOX) catalysis are open to a detailed structural analysis. Polyunsaturated fatty acids with two to six double bonds are oxygenated precisely on a particular carbon, typically forming a single chiral fatty acid hydroperoxide product. Molecular oxygen is not bound or liganded during catalysis, yet it is directed precisely to one position and one stereo configuration on the reacting fatty acid. The transformations proceed upon exposure of substrate to enzyme in the presence of O2 (RH + O2 → ROOH), so it has proved challenging to capture the precise mode of substrate binding in the LOX active site. Beginning with crystal structures with bound inhibitors or surrogate substrates, and most recently arachidonic acid bound under anaerobic conditions, a picture is consolidating of catalysis in a U-shaped fatty acid binding channel in which individual LOX enzymes use distinct amino acids to control the head-to-tail orientation of the fatty acid and register of the selected pentadiene opposite the non-heme iron, suitably positioned for the initial stereoselective hydrogen abstraction and subsequent reaction with O2 . Drawing on the crystal structures available currently, this review features the roles of the N-terminal β-barrel (C2-like, or PLAT domain) in substrate acquisition and sensitivity to cellular calcium, and the α-helical catalytic domain in fatty acid binding and reactions with O2 that produce hydroperoxide products with regio and stereospecificity. LOX structures combine to explain how similar enzymes with conserved catalytic machinery differ in product, but not substrate, specificities.
Keywords: arachidonic acid; hydroperoxide oxygenation; linoleic acid; lipoxygenase; oxylipins; polyunsaturated fatty acids.
© 2014 The Protein Society.
Figures
References
-
- Ivanov I, Heydeck D, Hofheinz K, Roffeis J, O'Donnell VB, Kuhn H, Walther M. Molecular enzymology of lipoxygenases. Arch Biochem Biophys. 2010;503:161–174. - PubMed
-
- Samuelsson B, Dahlen SE, Lindgren JA, Rouzer CA, Serhan CN. Leukotrienes and lipoxins: structures, biosynthesis, and biological effects. Science. 1987;237:1171–1176. - PubMed
-
- Andreou A, Feussner I. Lipoxygenases—structure and reaction mechanism. Phytochemistry. 2009;70:1504–1510. - PubMed
-
- Zheng Y, Yin H, Boeglin WE, Elias PM, Crumrine D, Beier DR, Brash AR. Lipoxygenases mediate the effect of essential fatty acid in skin barrier formation: a proposed role in releasing omega-hydroxyceramide for construction of the corneocyte lipid envelope. J Biol Chem. 2011;286:24046–24056. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
