

Postsynaptic P2X3-containing receptors in gustatory nerve fibres mediate responses to all taste qualities in mice
- PMID: 25524179
- PMCID: PMC4358674
- DOI: 10.1113/jphysiol.2014.281014
Postsynaptic P2X3-containing receptors in gustatory nerve fibres mediate responses to all taste qualities in mice
Abstract
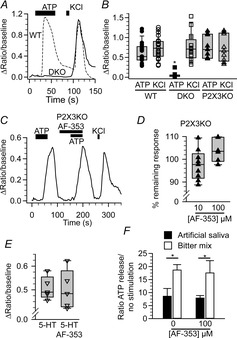
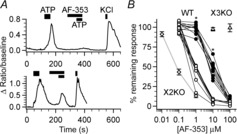
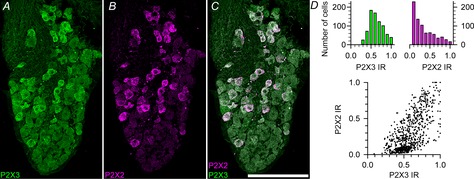
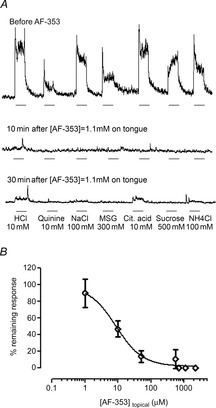
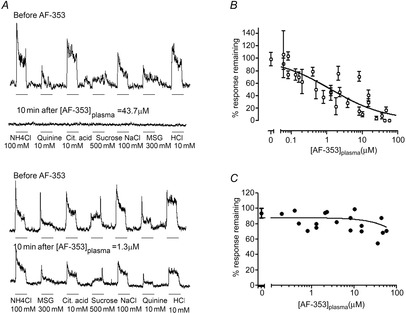
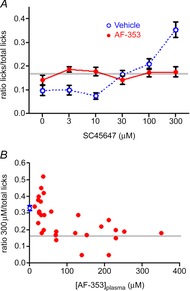
Taste buds release ATP to activate ionotropic purinoceptors composed of P2X2 and P2X3 subunits, present on the taste nerves. Mice with genetic deletion of P2X2 and P2X3 receptors (double knockout mice) lack responses to all taste stimuli presumably due to the absence of ATP-gated receptors on the afferent nerves. Recent experiments on the double knockout mice showed, however, that their taste buds fail to release ATP, suggesting the possibility of pleiotropic deficits in these global knockouts. To test further the role of postsynaptic P2X receptors in afferent signalling, we used AF-353, a selective antagonist of P2X3-containing receptors to inhibit the receptors acutely during taste nerve recording and behaviour. The specificity of AF-353 for P2X3-containing receptors was tested by recording Ca(2+) transients to exogenously applied ATP in fura-2 loaded isolated geniculate ganglion neurons from wild-type and P2X3 knockout mice. ATP responses were completely inhibited by 10 μm or 100 μm AF-353, but neither concentration blocked responses in P2X3 single knockout mice wherein the ganglion cells express only P2X2-containing receptors. Furthermore, AF-353 had no effect on taste-evoked ATP release from taste buds. In wild-type mice, i.p. injection of AF-353 or simple application of the drug directly to the tongue, inhibited taste nerve responses to all taste qualities in a dose-dependent fashion. A brief access behavioural assay confirmed the electrophysiological results and showed that preference for a synthetic sweetener, SC-45647, was abolished following i.p. injection of AF-353. These data indicate that activation of P2X3-containing receptors is required for transmission of all taste qualities.
© 2015 The Authors. The Journal of Physiology © 2015 The Physiological Society.
Figures
References
-
- Bo X, Alavi A, Xiang Z, Oglesby I, Ford A. Burnstock G. Localization of ATP-gated P2X2 and P2X3 receptor immunoreactive nerves in rat taste buds. Neuroreport. 1999;10:1107–1111. - PubMed
-
- Bystrova MF, Yatzenko YE, Fedorov IV, Rogachevskaja OA. Kolesnikov SS. P2Y isoforms operative in mouse taste cells. Cell Tissue Res. 2006;323:377–382. - PubMed
-
- Cao Y, Shreffler C. Herness S. Localization and functional investigation of the transcription factor CREB in taste receptor cells. Neuroreport. 2002;13:1321–1325. - PubMed
-
- Carter DS, Alam M, Cai H, Dillon MP, Ford AP, Gever JR, Jahangir A, Lin C, Moore AG, Wagner PJ. Zhai Y. Identification and SAR of novel diaminopyrimidines. Part1: The discovery of RO-4, a dual P2X(3)/P2X(2/3) antagonist for the treatment of pain. Bioorganic Med Chem Lett. 2009;19:1628–1631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous