

Transcriptional profiling at whole population and single cell levels reveals somatosensory neuron molecular diversity
- PMID: 25525749
- PMCID: PMC4383053
- DOI: 10.7554/eLife.04660
Transcriptional profiling at whole population and single cell levels reveals somatosensory neuron molecular diversity
Erratum in
-
Correction: Transcriptional profiling at whole population and single cell levels reveals somatosensory neuron molecular diversity.Elife. 2015 Feb 6;4:e06720. doi: 10.7554/eLife.06720. Elife. 2015. PMID: 25658835 Free PMC article. No abstract available.
Abstract
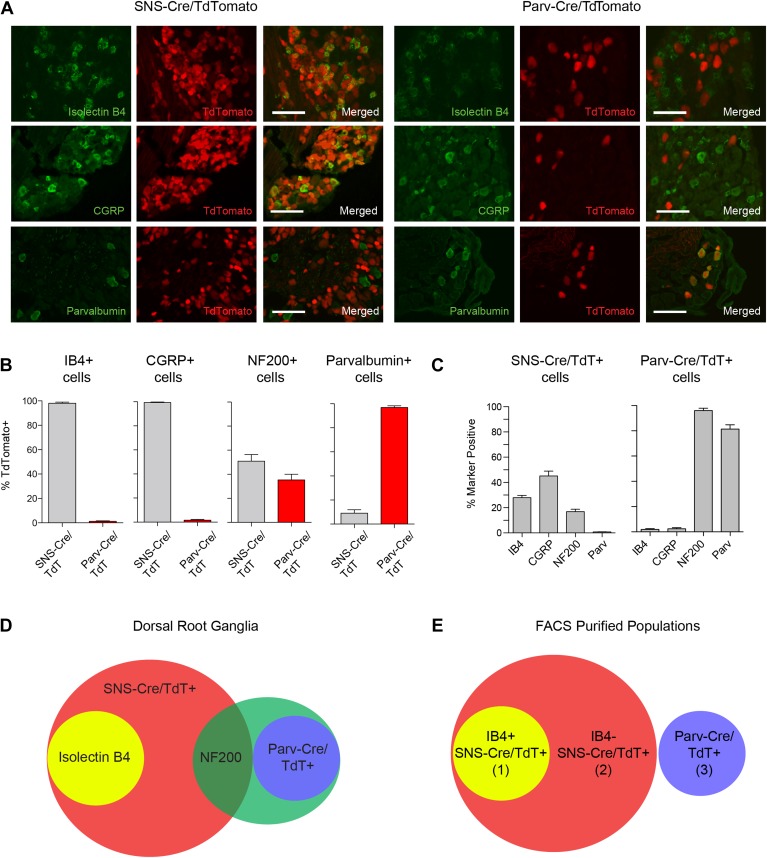
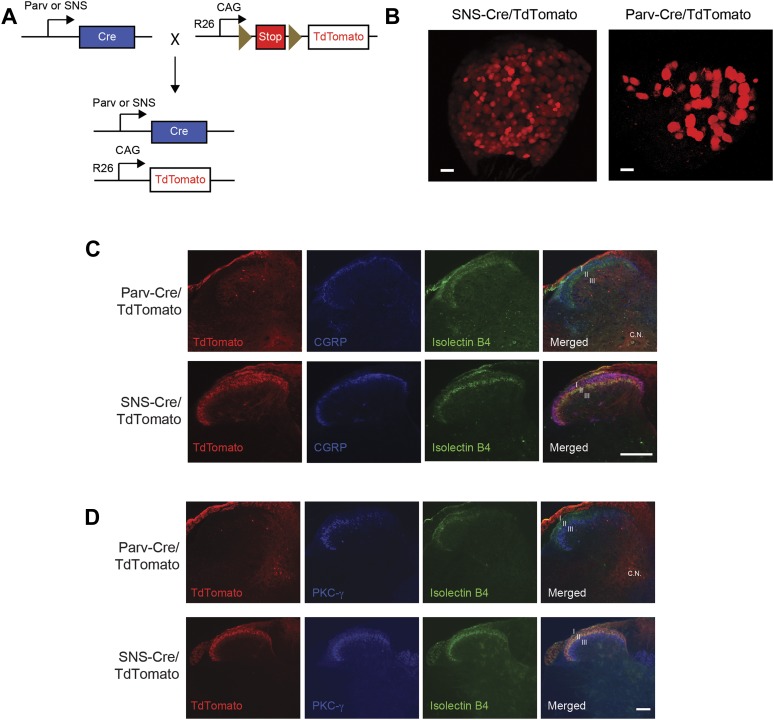
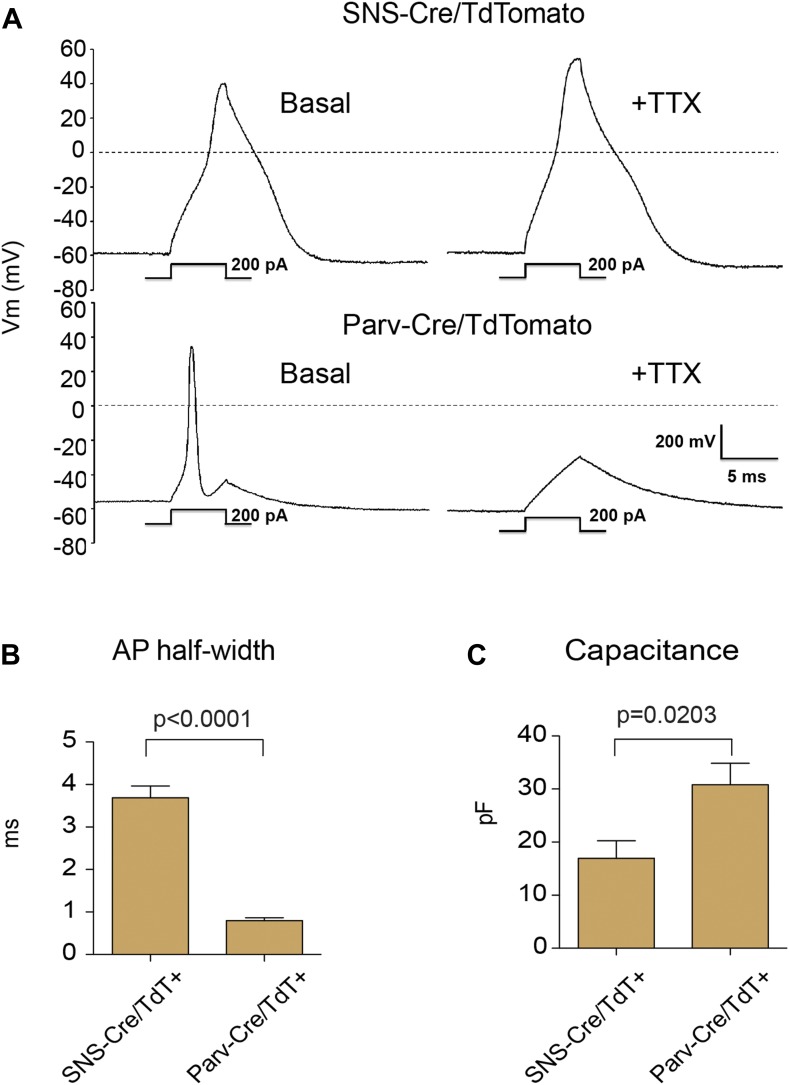
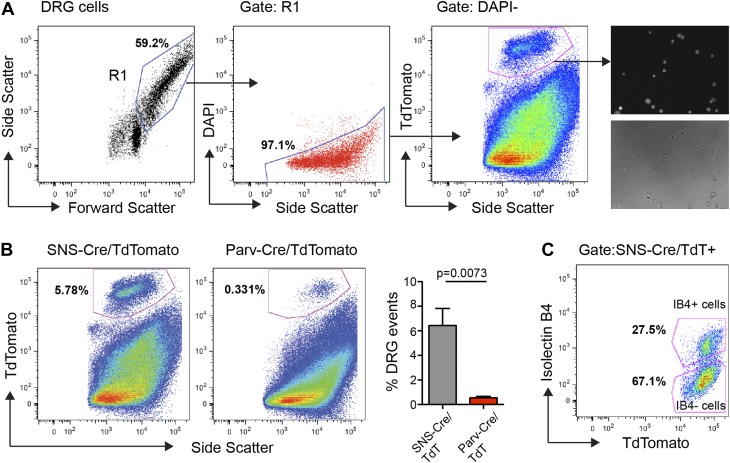
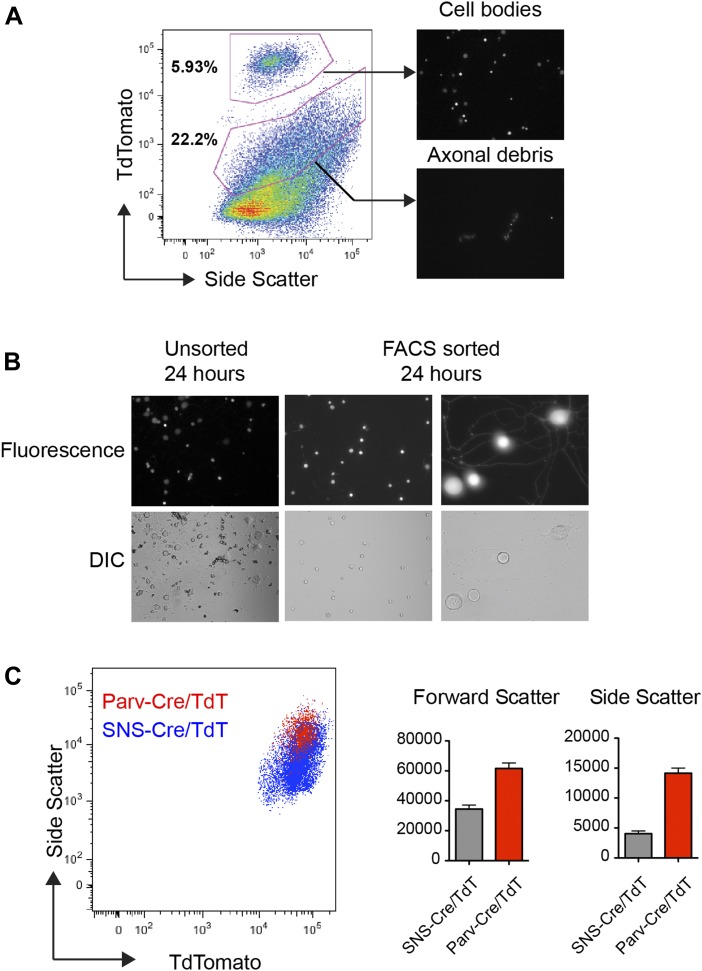
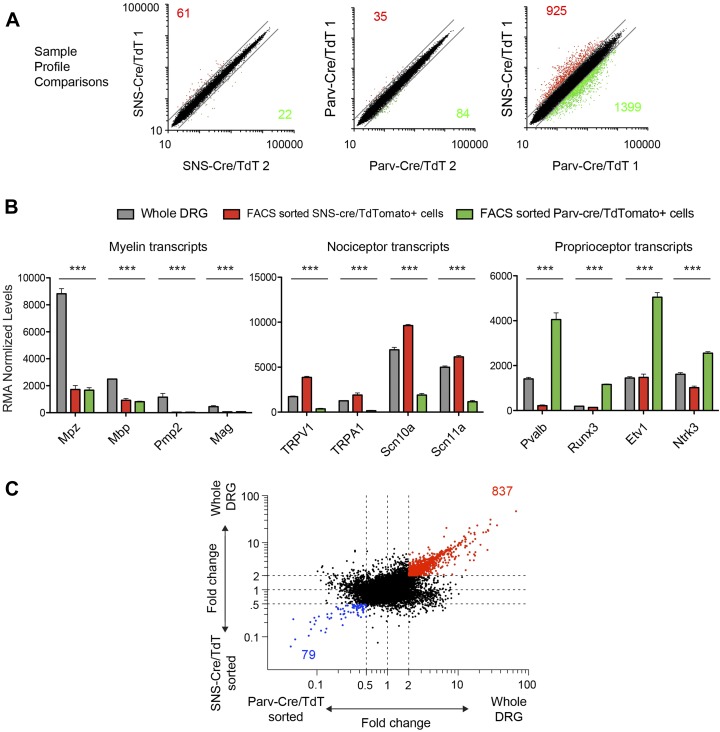
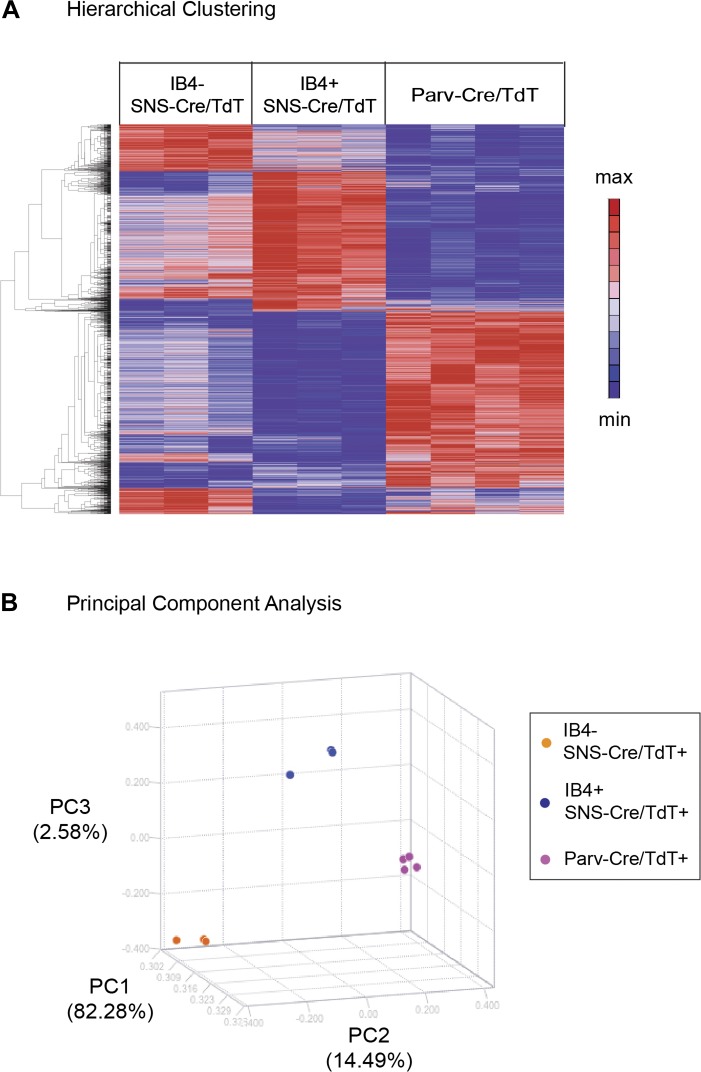
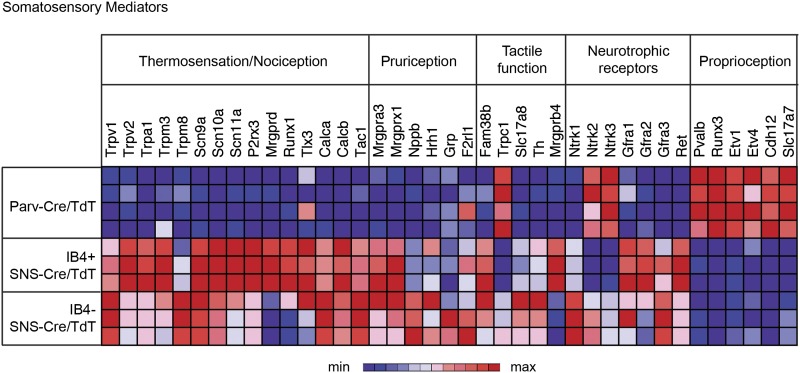
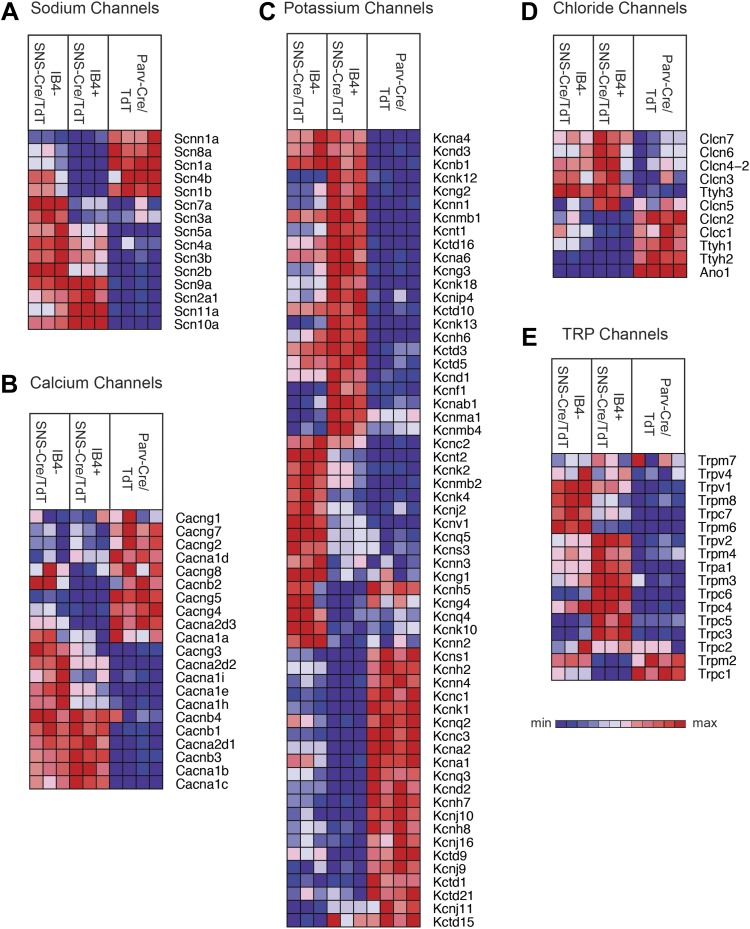
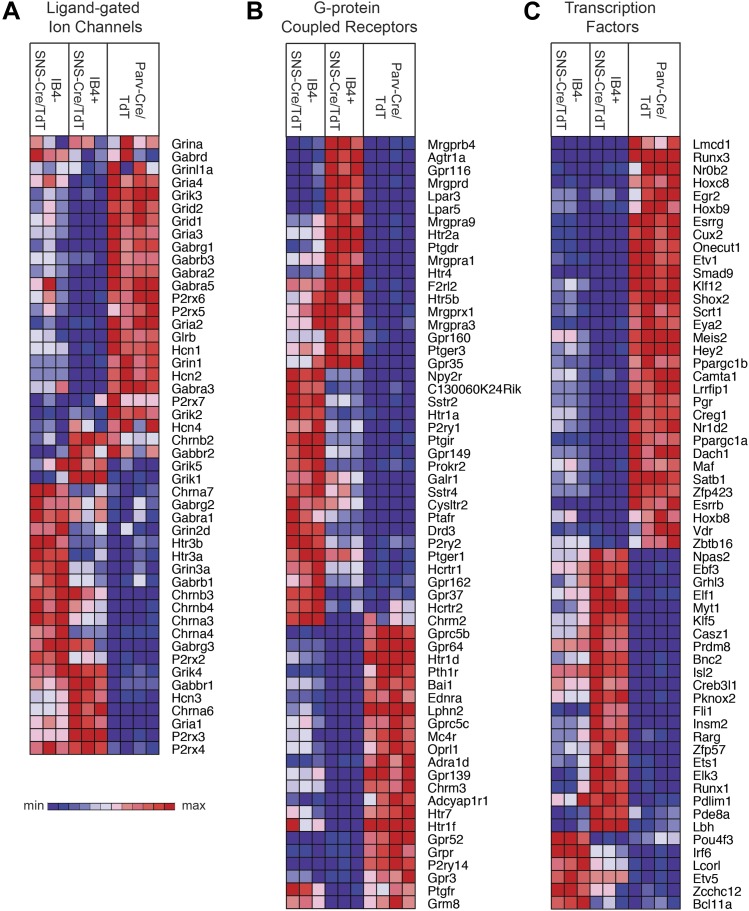
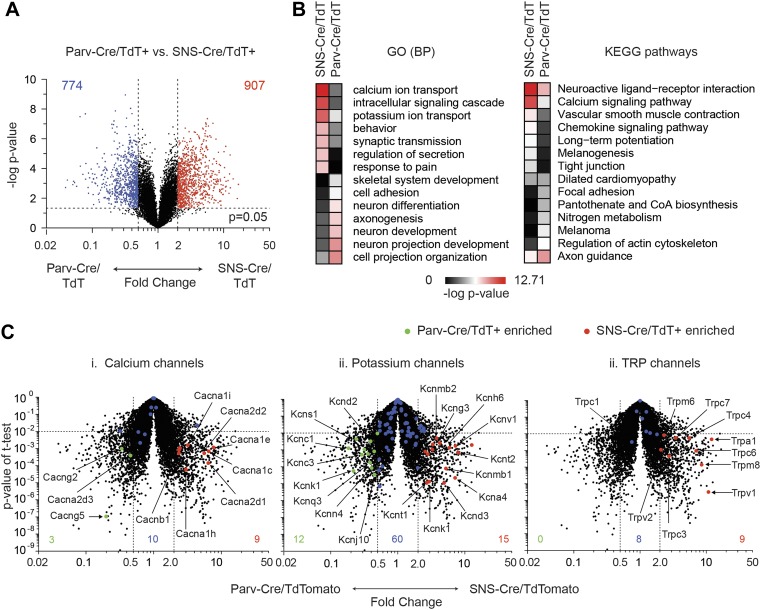
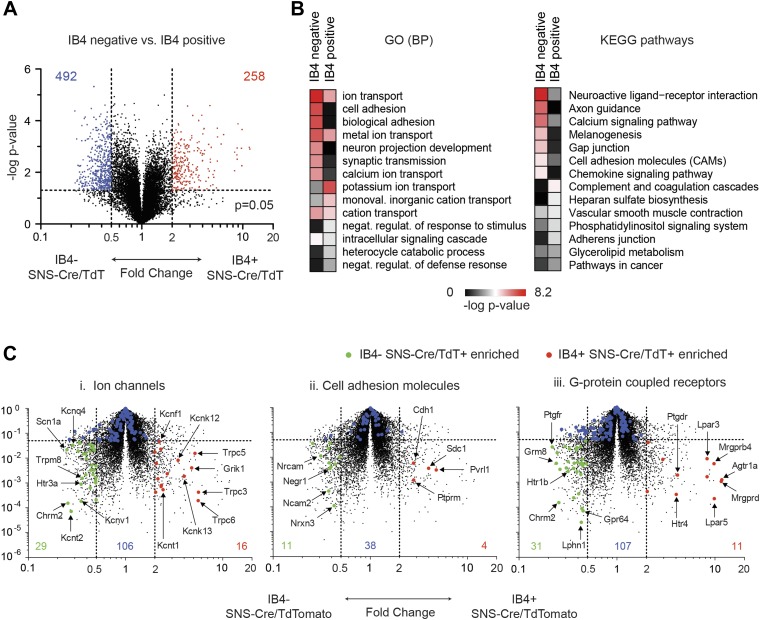
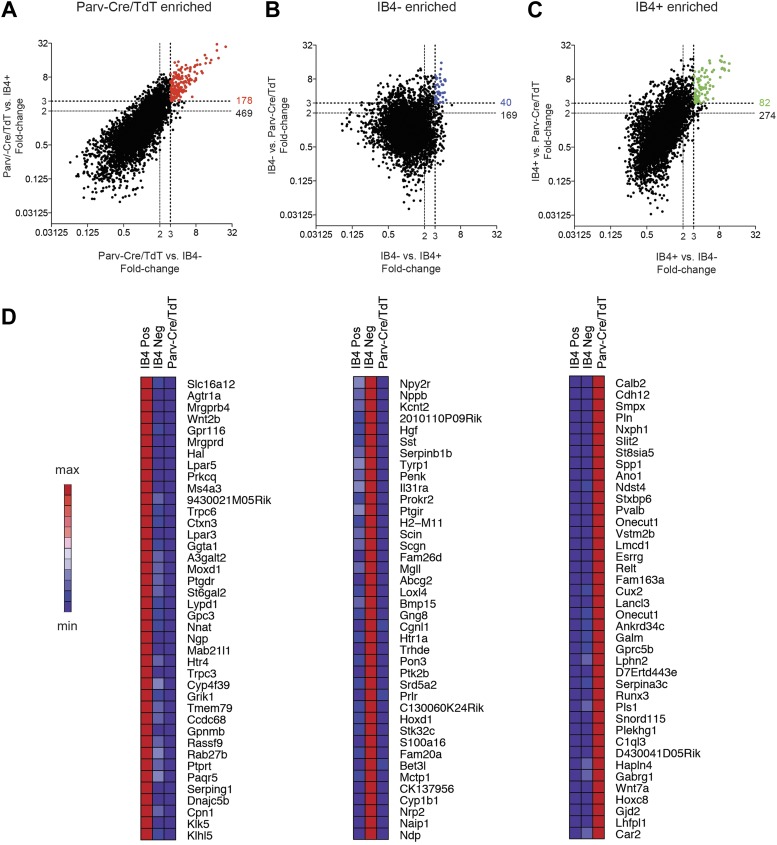
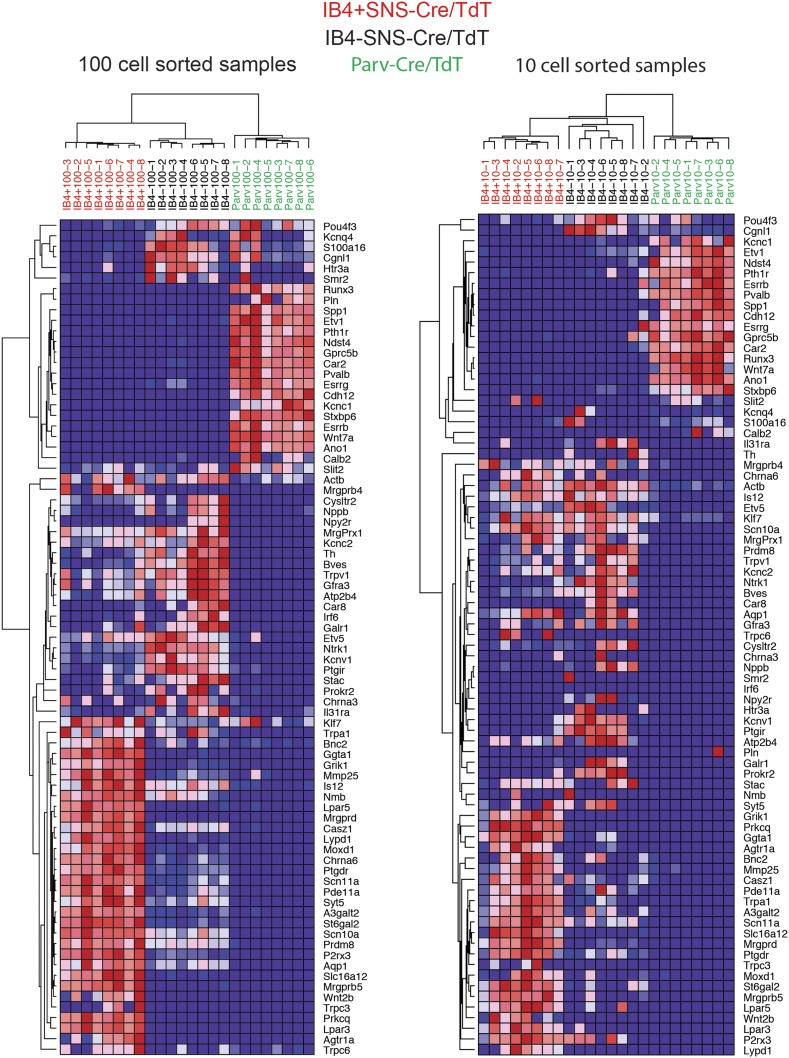
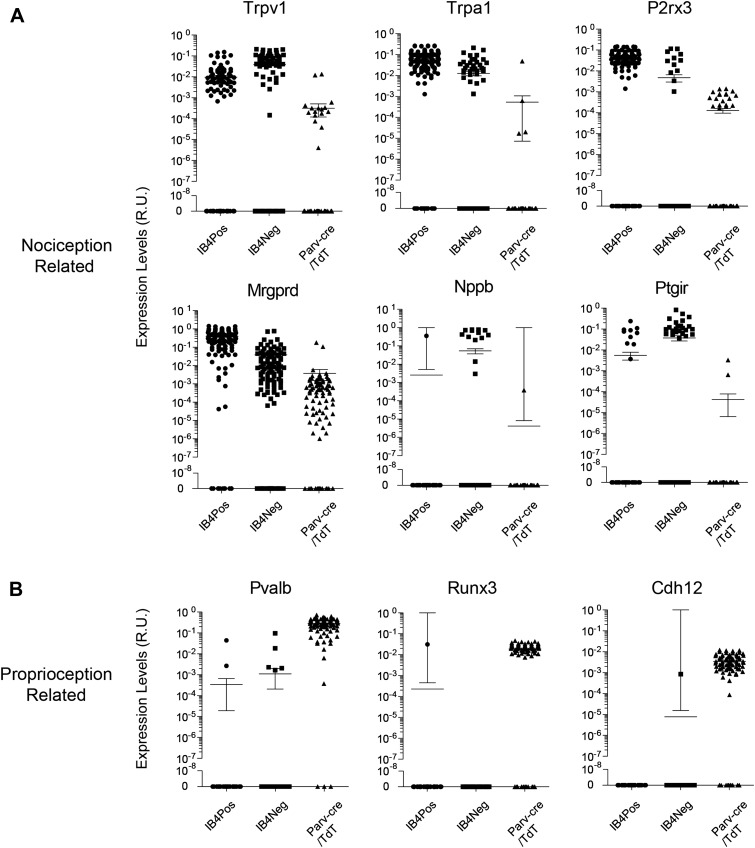
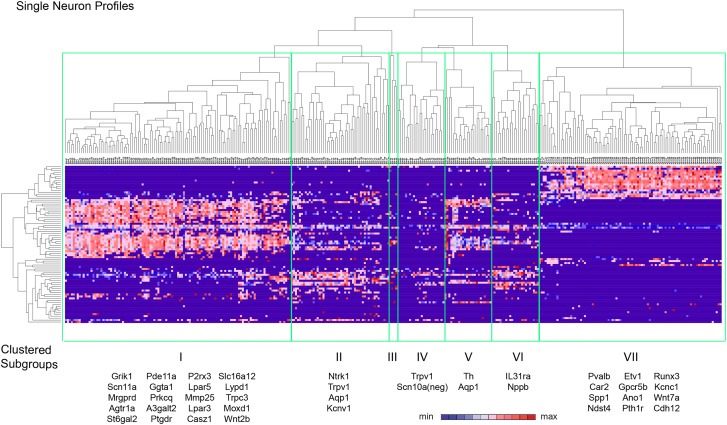
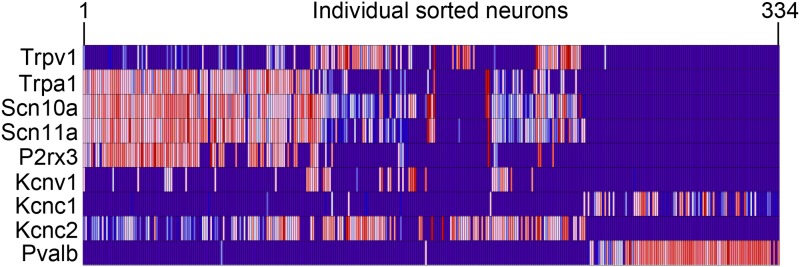
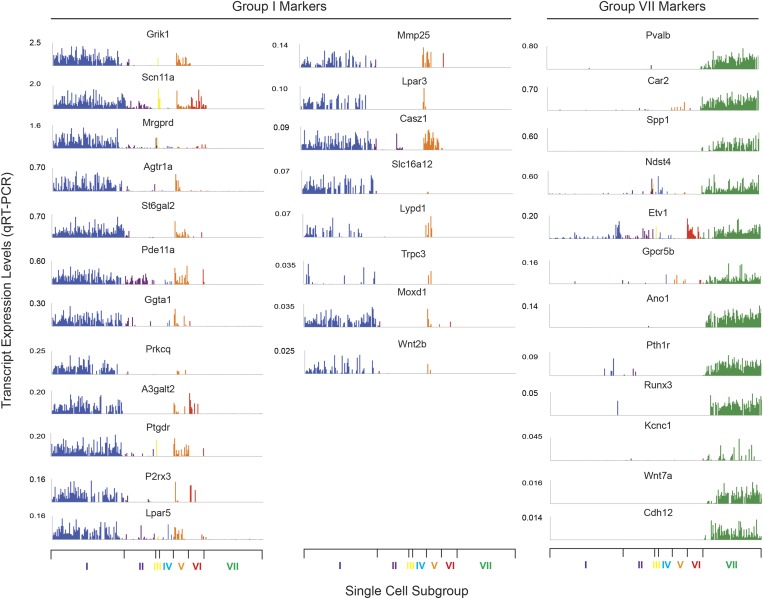
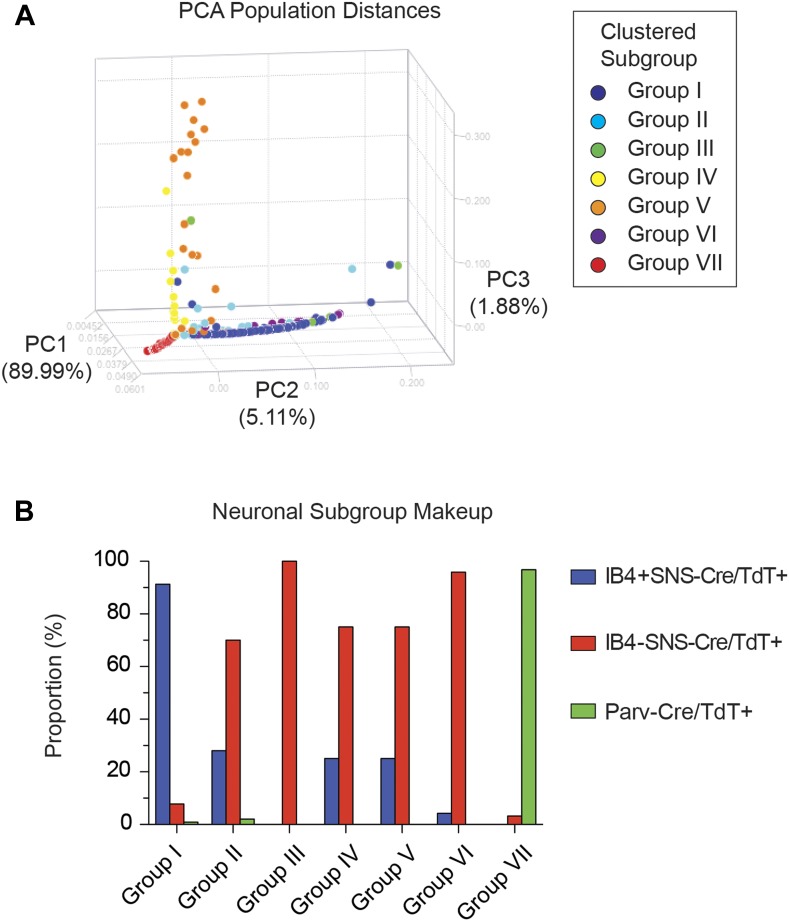
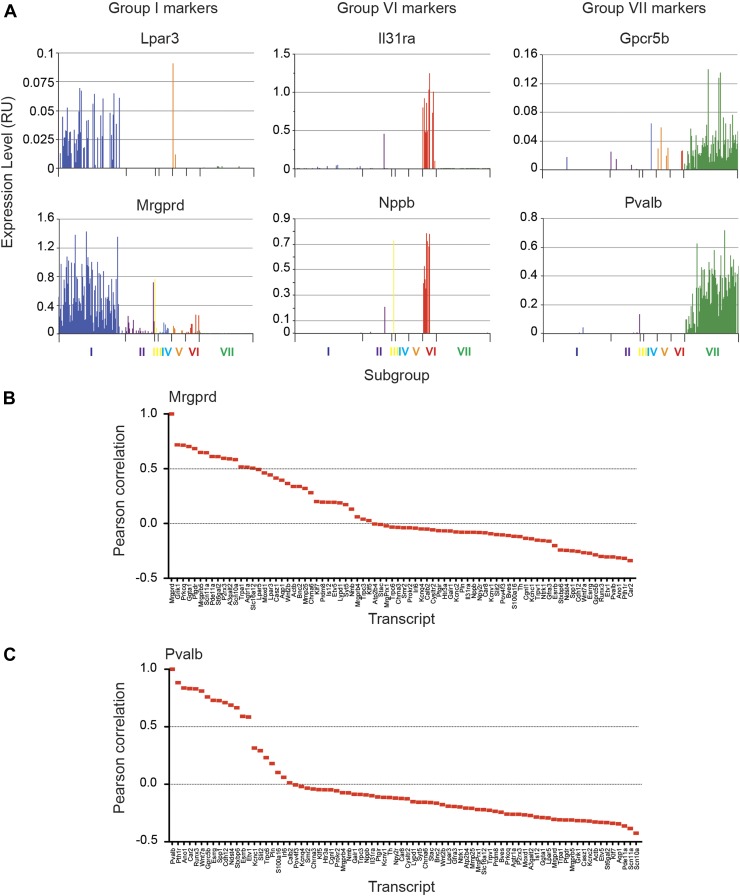
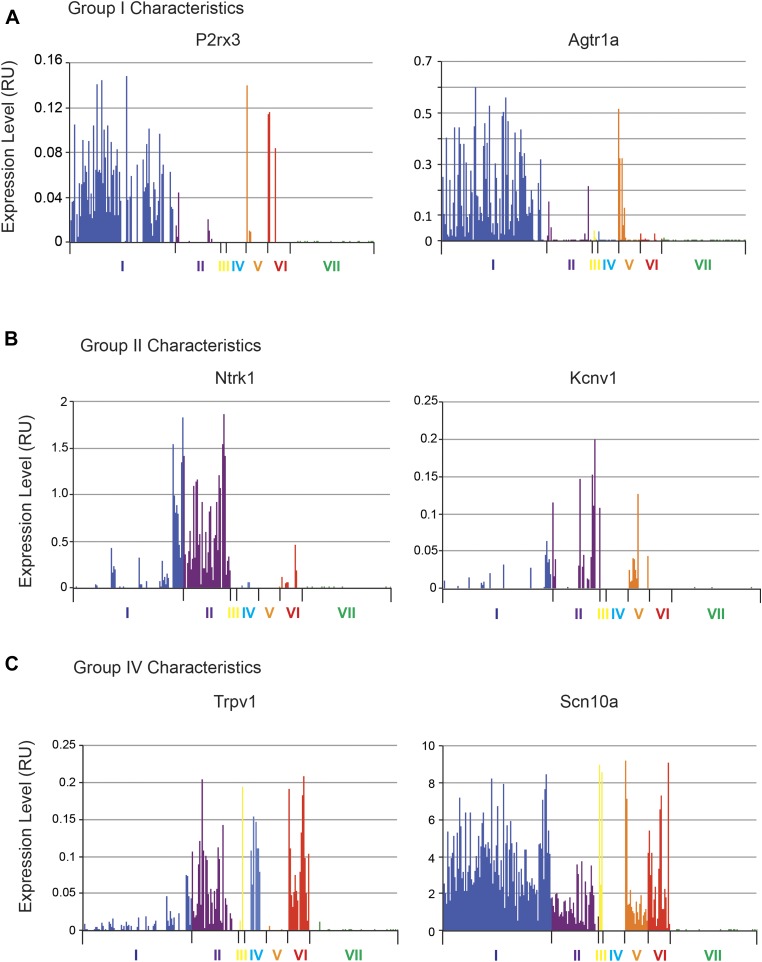
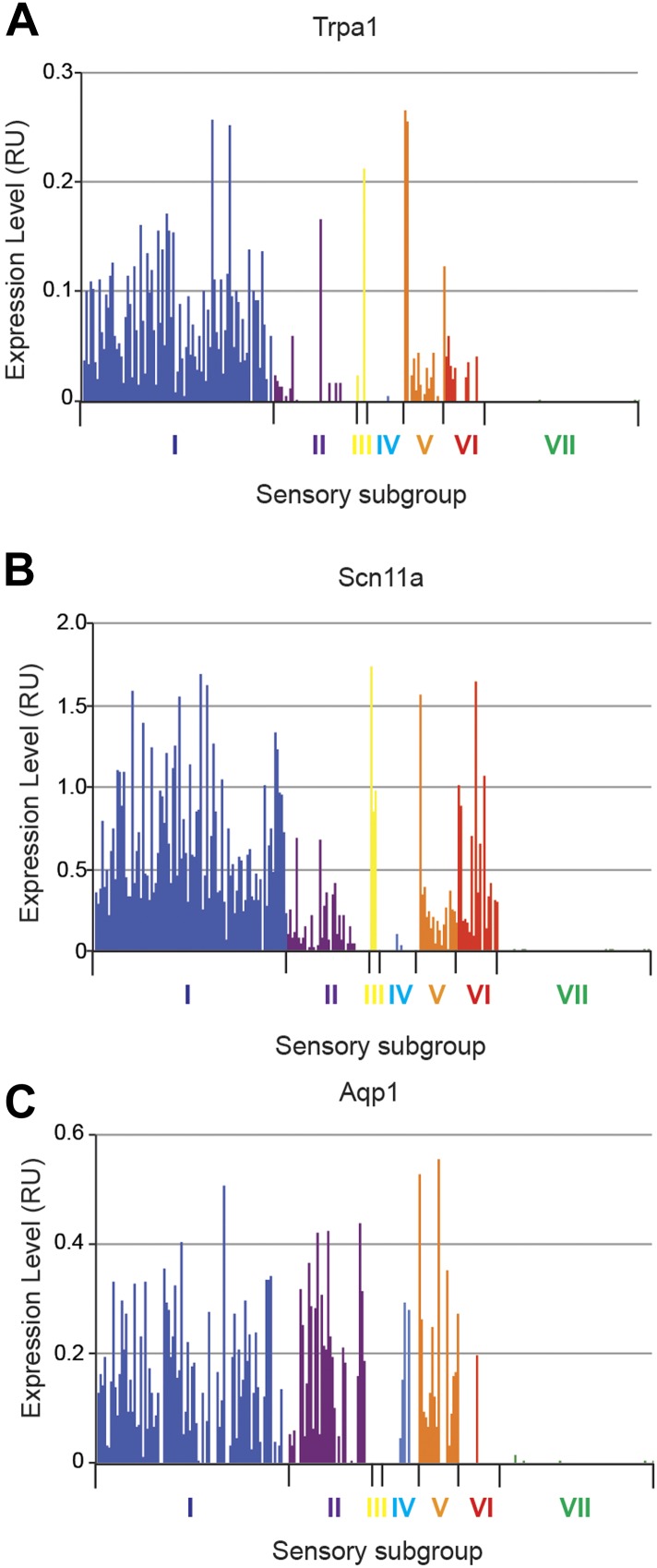
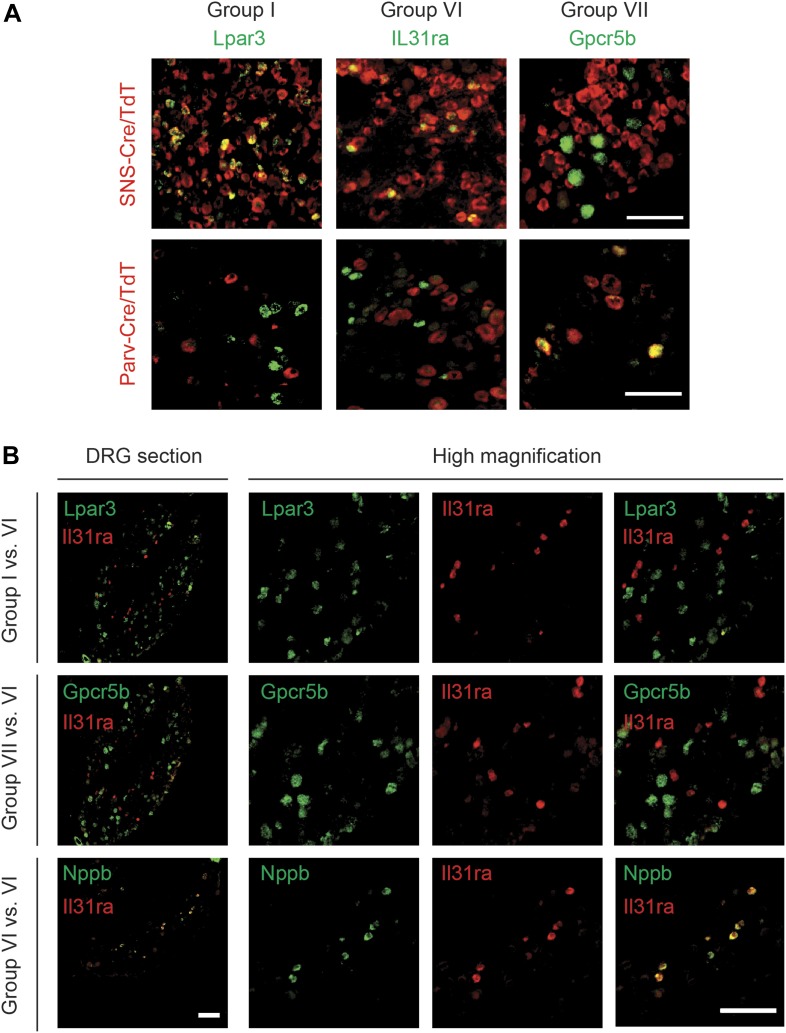
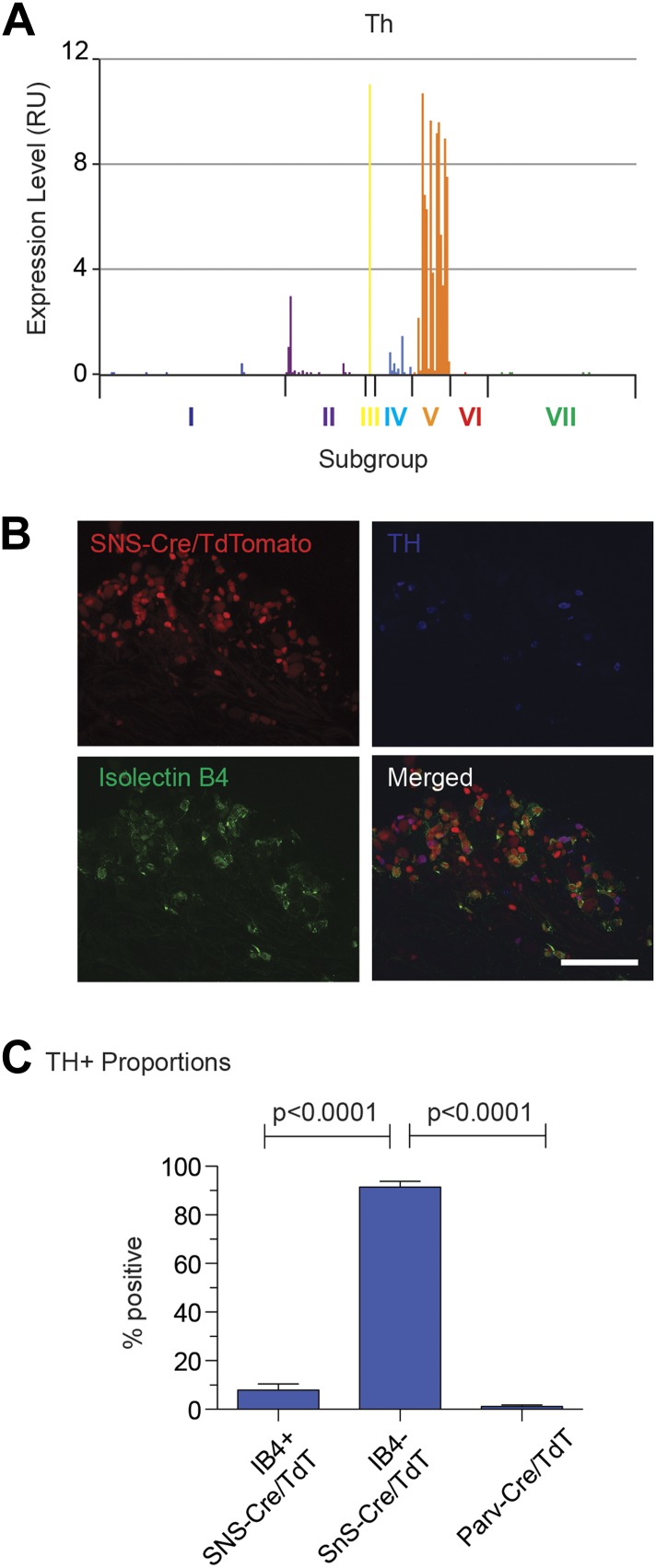
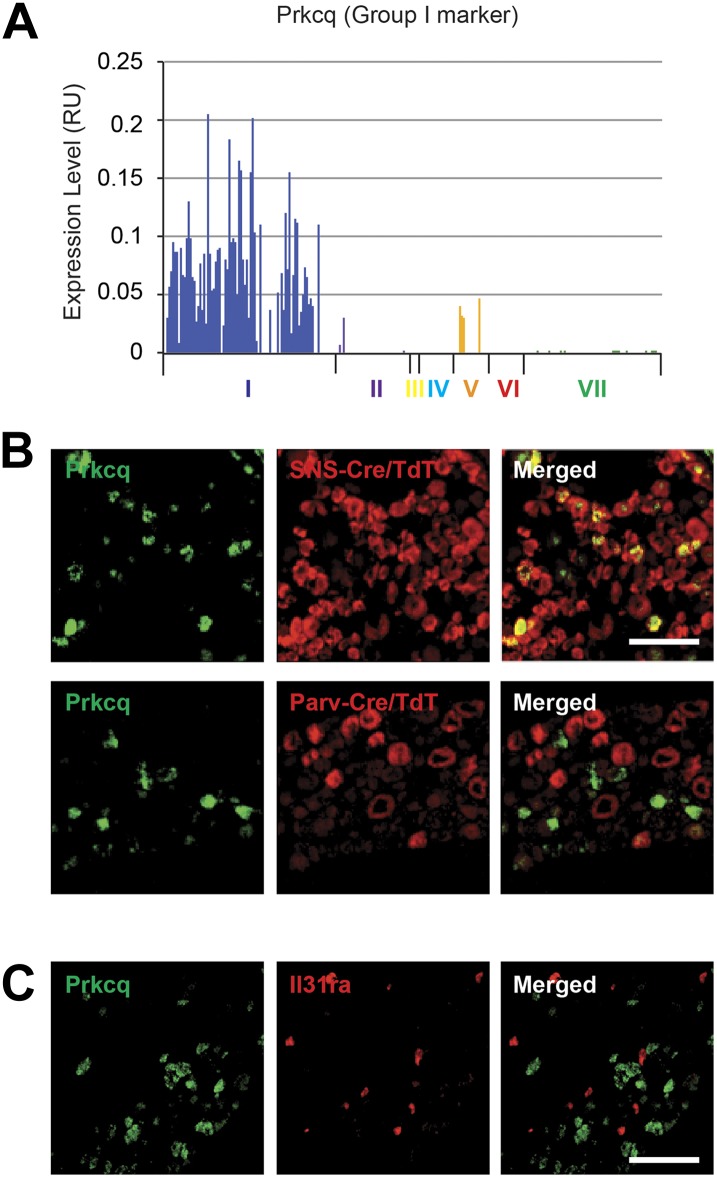
The somatosensory nervous system is critical for the organism's ability to respond to mechanical, thermal, and nociceptive stimuli. Somatosensory neurons are functionally and anatomically diverse but their molecular profiles are not well-defined. Here, we used transcriptional profiling to analyze the detailed molecular signatures of dorsal root ganglion (DRG) sensory neurons. We used two mouse reporter lines and surface IB4 labeling to purify three major non-overlapping classes of neurons: 1) IB4(+)SNS-Cre/TdTomato(+), 2) IB4(-)SNS-Cre/TdTomato(+), and 3) Parv-Cre/TdTomato(+) cells, encompassing the majority of nociceptive, pruriceptive, and proprioceptive neurons. These neurons displayed distinct expression patterns of ion channels, transcription factors, and GPCRs. Highly parallel qRT-PCR analysis of 334 single neurons selected by membership of the three populations demonstrated further diversity, with unbiased clustering analysis identifying six distinct subgroups. These data significantly increase our knowledge of the molecular identities of known DRG populations and uncover potentially novel subsets, revealing the complexity and diversity of those neurons underlying somatosensation.
Keywords: DRG; evolutionary biology; genomics; mouse; neuroscience; nociception; peripheral nervous system; proprioception; somatosensation; transcriptome.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
