

Hepatitis C virus RNA replication and assembly: living on the fat of the land
- PMID: 25525790
- PMCID: PMC7172941
- DOI: 10.1016/j.chom.2014.10.008
Hepatitis C virus RNA replication and assembly: living on the fat of the land
Abstract
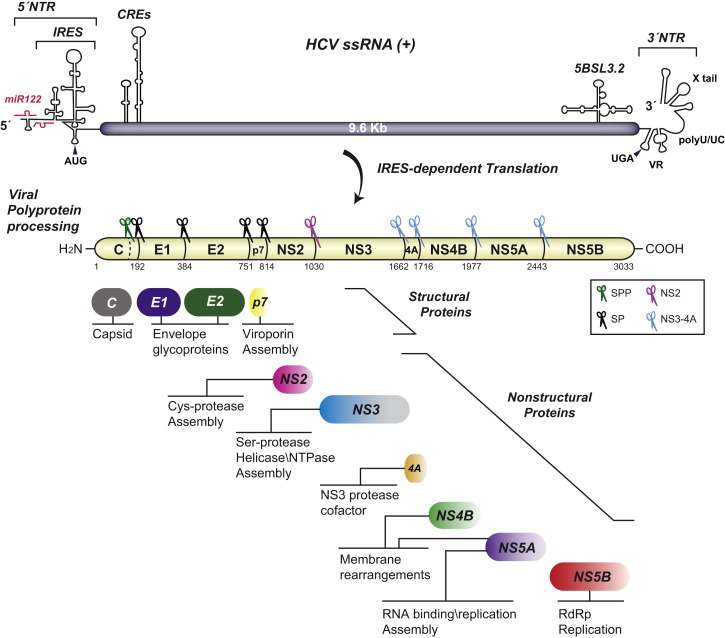
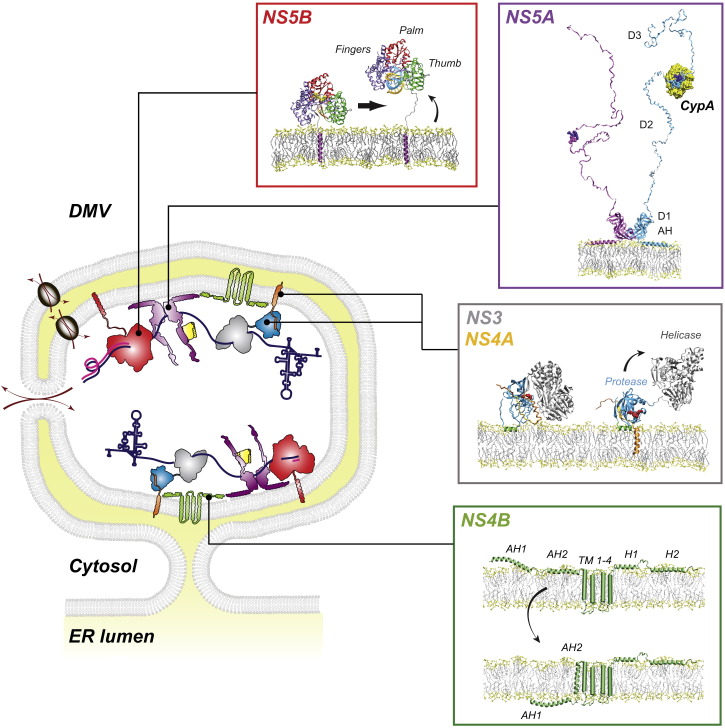
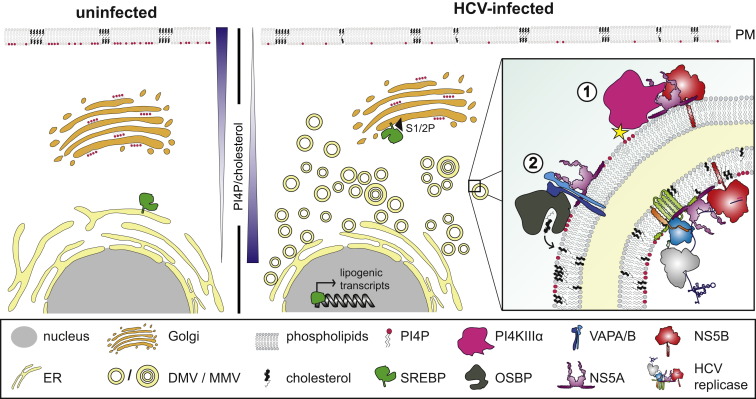
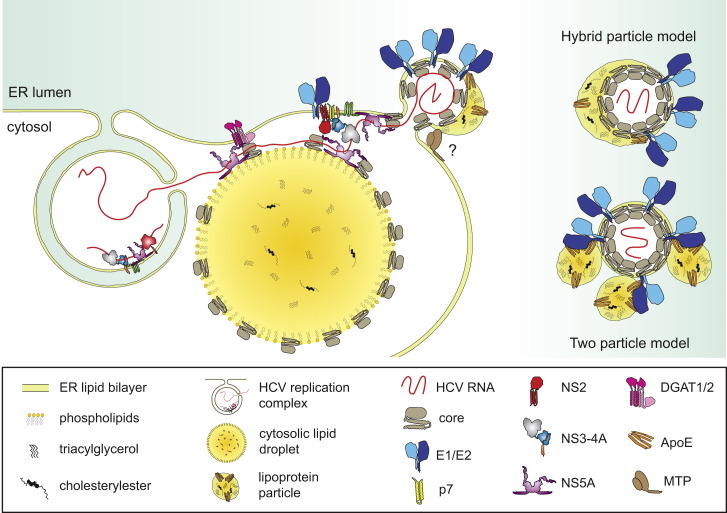
Hepatitis C virus (HCV) is a major global health burden accounting for around 170 million chronic infections worldwide. Although highly potent direct-acting antiviral drugs to treat chronic hepatitis C have been approved recently, owing to their high costs and limited availability and a large number of undiagnosed infections, the burden of disease is expected to rise in the next few years. In addition, HCV is an excellent paradigm for understanding the tight link between a pathogen and host cell pathways, most notably lipid metabolism. HCV extensively remodels intracellular membranes to establish its cytoplasmic replication factory and also usurps components of the intercellular lipid transport system for production of infectious virus particles. Here, we review the molecular mechanisms of viral replicase function, cellular pathways employed during HCV replication factory biogenesis, and viral, as well as cellular, determinants of progeny virus production.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Ago H., Adachi T., Yoshida A., Yamamoto M., Habuka N., Yatsunami K., Miyano M. Crystal structure of the RNA-dependent RNA polymerase of hepatitis C virus. Structure. 1999;7:1417–1426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
