

Regulation of tissue morphogenesis by endothelial cell-derived signals
- PMID: 25529933
- PMCID: PMC4943524
- DOI: 10.1016/j.tcb.2014.11.007
Regulation of tissue morphogenesis by endothelial cell-derived signals
Abstract
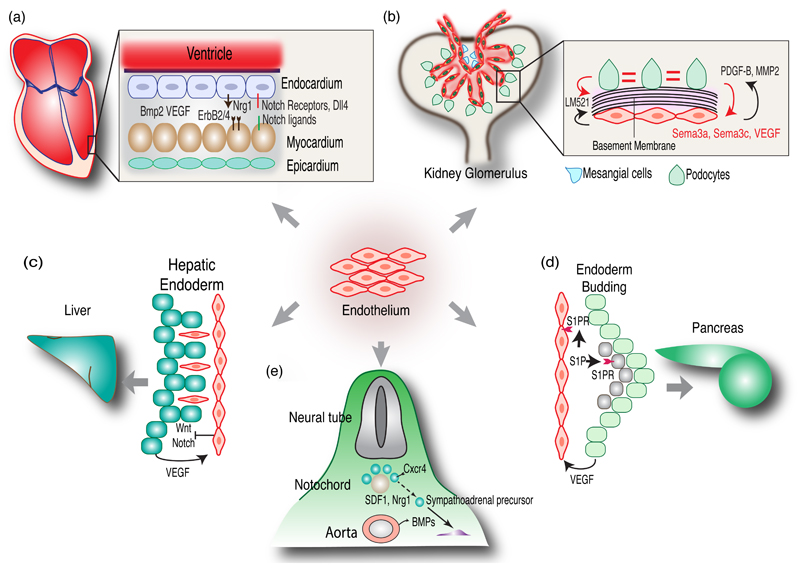
Endothelial cells (ECs) form an extensive network of blood vessels that has numerous essential functions in the vertebrate body. In addition to their well-established role as a versatile transport network, blood vessels can induce organ formation or direct growth and differentiation processes by providing signals in a paracrine (angiocrine) fashion. Tissue repair also requires the local restoration of vasculature. ECs are emerging as important signaling centers that coordinate regeneration and help to prevent deregulated, disease-promoting processes. Vascular cells are also part of stem cell niches and have key roles in hematopoiesis, bone formation, and neurogenesis. Here, we review these newly identified roles of ECs in the regulation of organ morphogenesis, maintenance, and regeneration.
Keywords: angiocrine signaling; angiogenesis; bone marrow; endothelial cells; liver; lung; organ morphogenesis; vascular niche.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures




References
-
- Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671–674. - PubMed
-
- Ehling M, et al. Notch controls retinal blood vessel maturation and quiescence. Development. 2013;140:3051–3061. - PubMed
-
- Korn C, et al. Endothelial cell-derived non-canonical Wnt ligands control vascular pruning in angiogenesis. Development. 2014;141:1757–1766. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
