

Redox metabolites signal polymicrobial biofilm development via the NapA oxidative stress cascade in Aspergillus
- PMID: 25532893
- PMCID: PMC4286458
- DOI: 10.1016/j.cub.2014.11.018
Redox metabolites signal polymicrobial biofilm development via the NapA oxidative stress cascade in Aspergillus
Abstract
Background: Filamentous fungi and bacteria form mixed-species biofilms in nature and diverse clinical contexts. They secrete a wealth of redox-active small molecule secondary metabolites, which are traditionally viewed as toxins that inhibit growth of competing microbes.
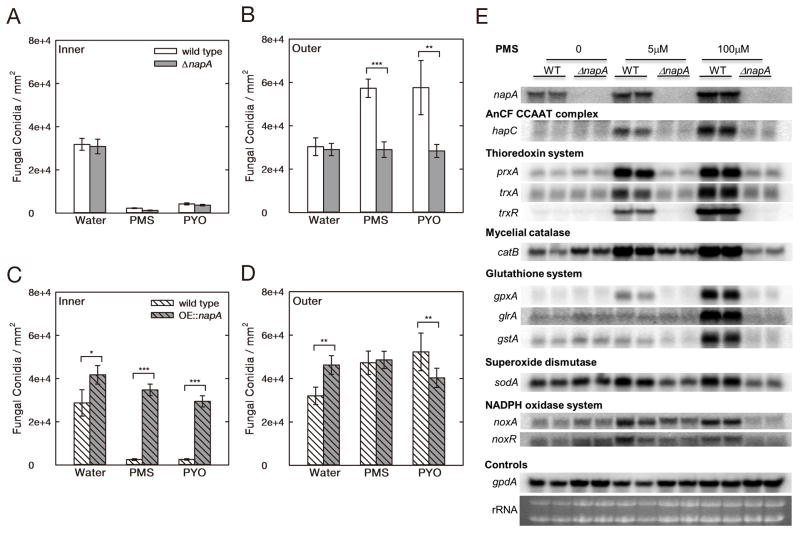
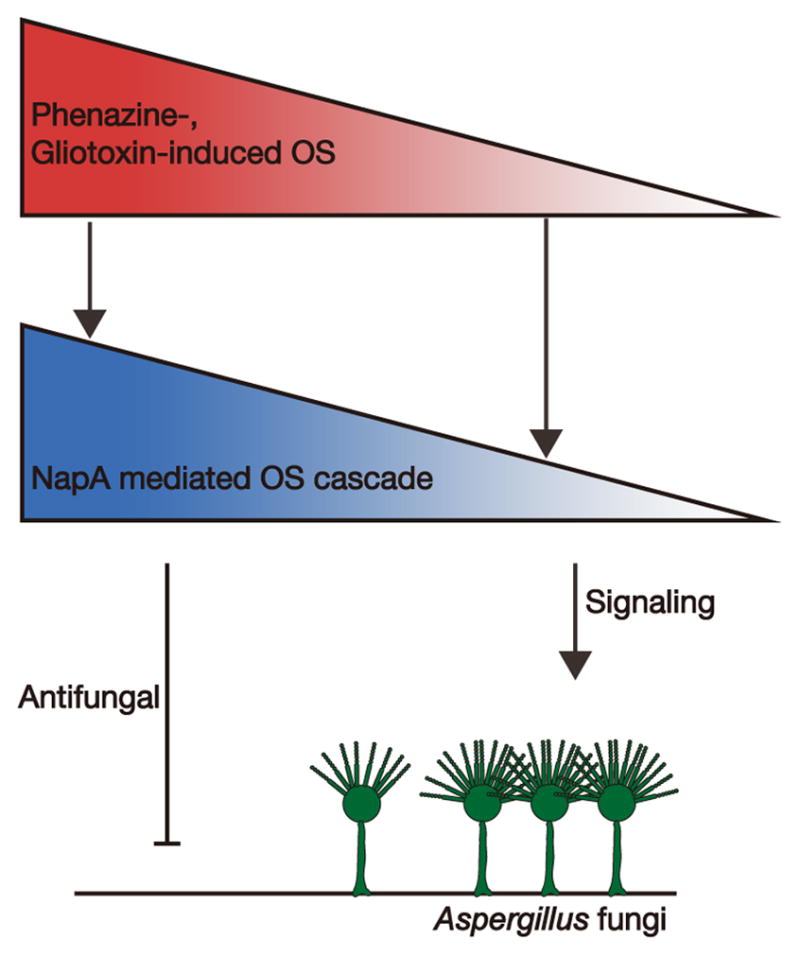
Results: Here, we report that these "toxins" can act as interspecies signals, affecting filamentous fungal development via oxidative stress regulation. Specifically, in coculture biofilms, Pseudomonas aeruginosa phenazine-derived metabolites differentially modulated Aspergillus fumigatus development, shifting from weak vegetative growth to induced asexual sporulation (conidiation) along a decreasing phenazine gradient. The A. fumigatus morphological shift correlated with the production of phenazine radicals and concomitant reactive oxygen species (ROS) production generated by phenazine redox cycling. Phenazine conidiation signaling was conserved in the genetic model A. nidulans and mediated by NapA, a homolog of AP-1-like bZIP transcription factor, which is essential for the response to oxidative stress in humans, yeast, and filamentous fungi. Expression profiling showed phenazine treatment induced a NapA-dependent response of the global oxidative stress metabolome, including the thioredoxin, glutathione, and NADPH-oxidase systems. Conidiation induction in A. nidulans by another microbial redox-active secondary metabolite, gliotoxin, also required NapA.
Conclusions: This work highlights that microbial redox metabolites are key signals for sporulation in filamentous fungi, which are communicated through an evolutionarily conserved eukaryotic stress response pathway. It provides a foundation for interspecies signaling in environmental and clinical biofilms involving bacteria and filamentous fungi.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures




Similar articles
-
Illumina identification of RsrA, a conserved C2H2 transcription factor coordinating the NapA mediated oxidative stress signaling pathway in Aspergillus.BMC Genomics. 2014 Nov 22;15(1):1011. doi: 10.1186/1471-2164-15-1011. BMC Genomics. 2014. PMID: 25416206 Free PMC article.
-
Secondary Metabolites Produced during Aspergillus fumigatus and Pseudomonas aeruginosa Biofilm Formation.mBio. 2022 Aug 30;13(4):e0185022. doi: 10.1128/mbio.01850-22. Epub 2022 Jul 20. mBio. 2022. PMID: 35856657 Free PMC article.
-
The Heterotrimeric Transcription Factor CCAAT-Binding Complex and Ca2+-CrzA Signaling Reversely Regulate the Transition between Fungal Hyphal Growth and Asexual Reproduction.mBio. 2021 Dec 21;12(6):e0300721. doi: 10.1128/mBio.03007-21. Epub 2021 Nov 16. mBio. 2021. PMID: 34781745 Free PMC article.
-
Resistance is not futile: gliotoxin biosynthesis, functionality and utility.Trends Microbiol. 2015 Jul;23(7):419-28. doi: 10.1016/j.tim.2015.02.005. Epub 2015 Mar 10. Trends Microbiol. 2015. PMID: 25766143 Review.
-
Genetic control of asexual development in aspergillus fumigatus.Adv Appl Microbiol. 2015;90:93-107. doi: 10.1016/bs.aambs.2014.09.004. Epub 2014 Nov 12. Adv Appl Microbiol. 2015. PMID: 25596030 Review.
Cited by
-
Beyond Penicillin: The Potential of Filamentous Fungi for Drug Discovery in the Age of Antibiotic Resistance.Antibiotics (Basel). 2023 Jul 29;12(8):1250. doi: 10.3390/antibiotics12081250. Antibiotics (Basel). 2023. PMID: 37627670 Free PMC article. Review.
-
Oxidative stress response pathways in fungi.Cell Mol Life Sci. 2022 Jun 1;79(6):333. doi: 10.1007/s00018-022-04353-8. Cell Mol Life Sci. 2022. PMID: 35648225 Free PMC article. Review.
-
Pseudomonas aeruginosa PumA acts on an endogenous phenazine to promote self-resistance.Microbiology (Reading). 2018 May;164(5):790-800. doi: 10.1099/mic.0.000657. Epub 2018 Apr 9. Microbiology (Reading). 2018. PMID: 29629858 Free PMC article.
-
Biofilm Filtrates of Pseudomonas aeruginosa Strains Isolated from Cystic Fibrosis Patients Inhibit Preformed Aspergillus fumigatus Biofilms via Apoptosis.PLoS One. 2016 Mar 1;11(3):e0150155. doi: 10.1371/journal.pone.0150155. eCollection 2016. PLoS One. 2016. PMID: 26930399 Free PMC article.
-
Establishing a Biofilm Co-culture of Pseudomonas and Aspergillus for Metabolite Extraction.Bio Protoc. 2015 Dec 5;5(23):e1667. doi: 10.21769/BioProtoc.1667. Bio Protoc. 2015. PMID: 29104889 Free PMC article.
References
-
- Amin R, Dupuis A, Aaron SD, Ratjen F. The effect of chronic infection with Aspergillus fumigatus on lung function and hospitalization in patients with cystic fibrosis. Chest. 2010;137:171–176. - PubMed
-
- Peleg AY, Hogan DA, Mylonakis E. Medically important bacterial-fungal interactions. Nat Rev Microbiol. 2010;8:340–349. - PubMed
-
- Costerton JW, Stewart PS, Creenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1999;284:1318–1323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
