

Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation
- PMID: 25533336
- PMCID: PMC4293269
- DOI: 10.1016/j.ccell.2014.11.009
Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation
Abstract
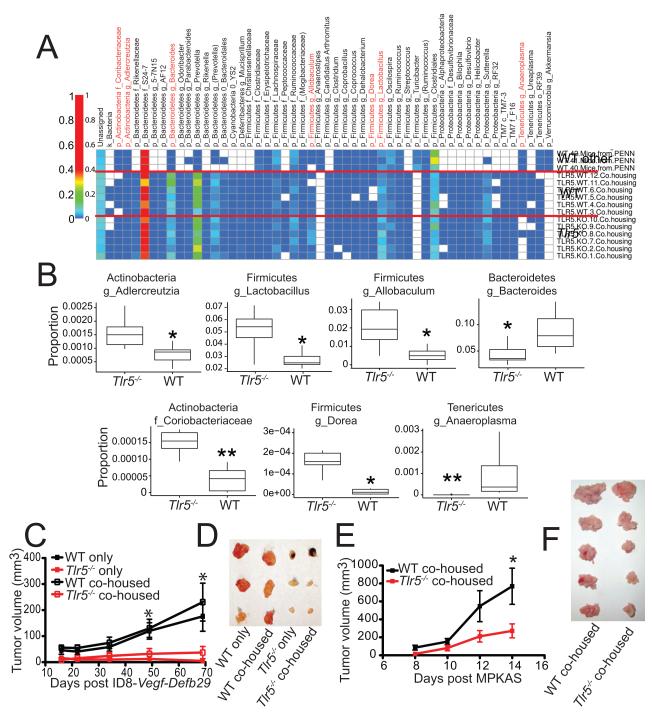
The dominant TLR5(R392X) polymorphism abrogates flagellin responses in >7% of humans. We report that TLR5-dependent commensal bacteria drive malignant progression at extramucosal locations by increasing systemic IL-6, which drives mobilization of myeloid-derived suppressor cells (MDSCs). Mechanistically, expanded granulocytic MDSCs cause γδ lymphocytes in TLR5-responsive tumors to secrete galectin-1, dampening antitumor immunity and accelerating malignant progression. In contrast, IL-17 is consistently upregulated in TLR5-unresponsive tumor-bearing mice but only accelerates malignant progression in IL-6-unresponsive tumors. Importantly, depletion of commensal bacteria abrogates TLR5-dependent differences in tumor growth. Contrasting differences in inflammatory cytokines and malignant evolution are recapitulated in TLR5-responsive/unresponsive ovarian and breast cancer patients. Therefore, inflammation, antitumor immunity, and the clinical outcome of cancer patients are influenced by a common TLR5 polymorphism.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures







Comment in
-
Common TLR5 mutations control cancer progression.Cancer Cell. 2015 Jan 12;27(1):1-3. doi: 10.1016/j.ccell.2014.12.008. Cancer Cell. 2015. PMID: 25584886
References
-
- Casanova JL, Abel L, Quintana-Murci L. Human TLRs and IL-1Rs in host defense: natural insights from evolutionary, epidemiological, and clinical genetics. Annu Rev Immunol. 2011;29:447–491. - PubMed
-
- Conejo-Garcia JR, Benencia F, Courreges MC, Kang E, Mohamed-Hadley A, Buckanovich RJ, Holtz DO, Jenkins A, Na H, Zhang L, et al. Tumor-infiltrating dendritic cell precursors recruited by a beta-defensin contribute to vasculogenesis under the influence of Vegf-A. Nat Med. 2004;10:950–958. - PubMed
-
- Cubillos-Ruiz J, Engle X, Scarlett U, Martinez D, Barber A, Elgueta R, Wang L, Nesbeth Y, Durant Y, Gewirtz A, et al. Polyethylenimine-based siRNA nanocomplexes reprogram tumor-associated dendritic cells via TLR5 to elicit therapeutic antitumor immunity. The Journal of clinical investigation. 2009;119:2231–2244. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- P30CA10815/CA/NCI NIH HHS/United States
- R01CA178687/CA/NCI NIH HHS/United States
- U54 CA151662/CA/NCI NIH HHS/United States
- T32CA009171/CA/NCI NIH HHS/United States
- R01CA157664/CA/NCI NIH HHS/United States
- P30 CA010815/CA/NCI NIH HHS/United States
- R01CA124515/CA/NCI NIH HHS/United States
- U54CA151662/CA/NCI NIH HHS/United States
- R01 CA124515/CA/NCI NIH HHS/United States
- CA010815/CA/NCI NIH HHS/United States
- T32 CA009171/CA/NCI NIH HHS/United States
- R01 CA163377/CA/NCI NIH HHS/United States
- R01 CA157664/CA/NCI NIH HHS/United States
- R01 CA178687/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
