

A tethered agonist within the ectodomain activates the adhesion G protein-coupled receptors GPR126 and GPR133
- PMID: 25533341
- PMCID: PMC4277498
- DOI: 10.1016/j.celrep.2014.11.036
A tethered agonist within the ectodomain activates the adhesion G protein-coupled receptors GPR126 and GPR133
Erratum in
-
A Tethered Agonist within the Ectodomain Activates the Adhesion G Protein-Coupled Receptors GPR126 and GPR133.Cell Rep. 2015 Feb 17;10(6):1021. doi: 10.1016/j.celrep.2015.01.065. Epub 2015 Feb 17. Cell Rep. 2015. PMID: 30849857 No abstract available.
Abstract
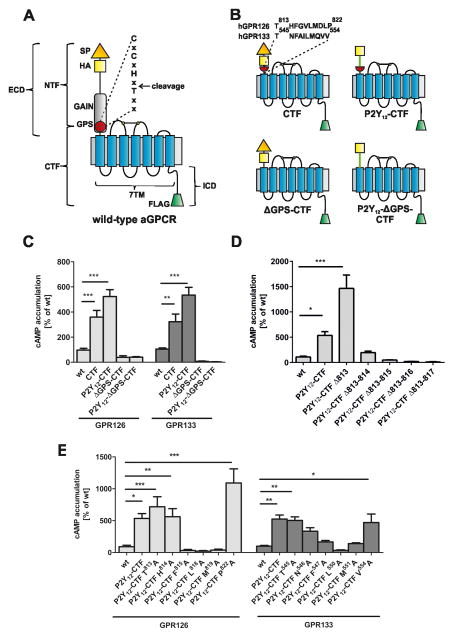
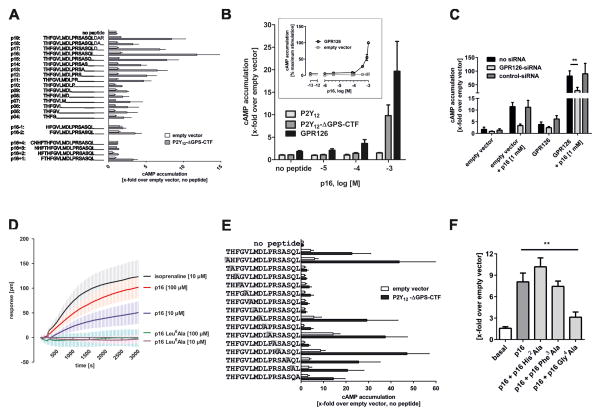
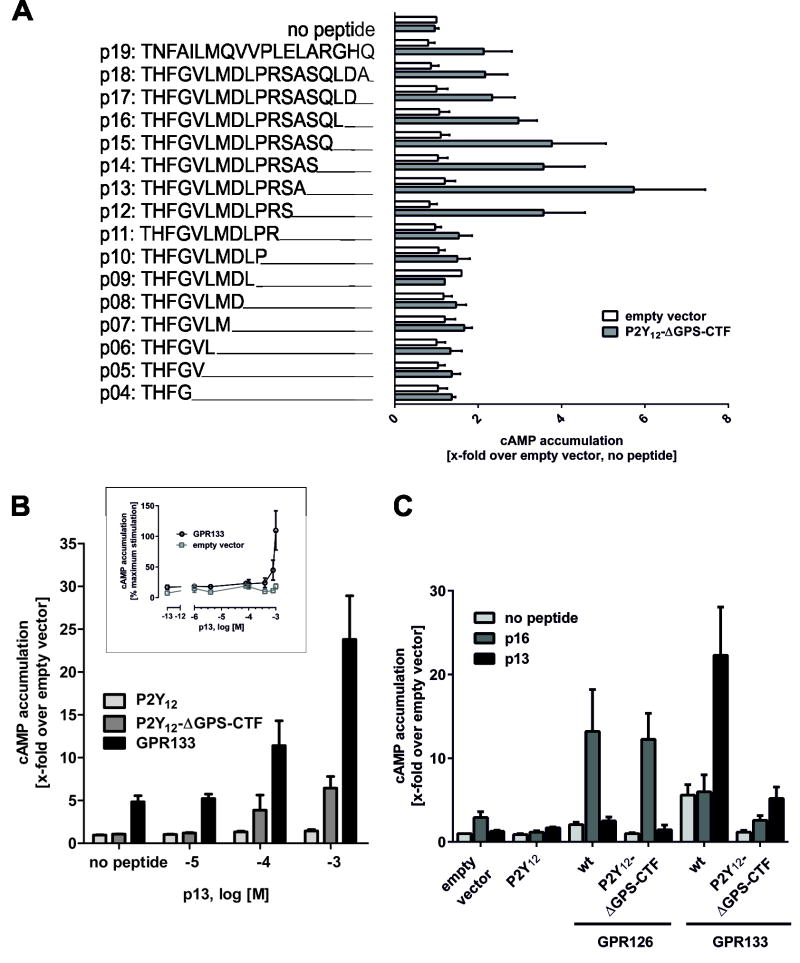
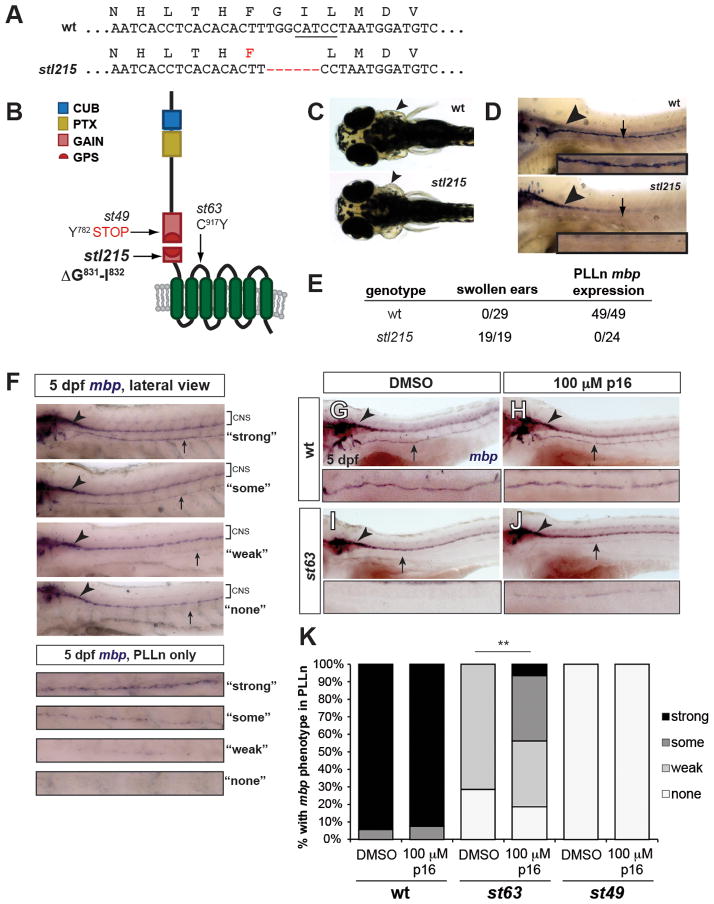
Adhesion G protein-coupled receptors (aGPCRs) comprise the second largest yet least studied class of the GPCR superfamily. aGPCRs are involved in many developmental processes and immune and synaptic functions, but the mode of their signal transduction is unclear. Here, we show that a short peptide sequence (termed the Stachel sequence) within the ectodomain of two aGPCRs (GPR126 and GPR133) functions as a tethered agonist. Upon structural changes within the receptor ectodomain, this intramolecular agonist is exposed to the seven-transmembrane helix domain, which triggers G protein activation. Our studies show high specificity of a given Stachel sequence for its receptor. Finally, the function of Gpr126 is abrogated in zebrafish with a mutated Stachel sequence, and signaling is restored in hypomorphic gpr126 zebrafish mutants upon exogenous Stachel peptide application. These findings illuminate a mode of aGPCR activation and may prompt the development of specific ligands for this currently untargeted GPCR family.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Bjarnadottir TK, Fredriksson R, Hoglund PJ, Gloriam DE, Lagerstrom MC, Schioth HB. The human and mouse repertoire of the adhesion family of G-protein-coupled receptors. Genomics. 2004;84:23–33. - PubMed
-
- Geng FS, Abbas L, Baxendale S, Holdsworth CJ, Swanson AG, Slanchev K, Hammerschmidt M, Topczewski J, Whitfield TT. Semicircular canal morphogenesis in the zebrafish inner ear requires the function of gpr126 (lauscher), an adhesion class G protein-coupled receptor gene. Development. 2013;140:4362–4374. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
