

TAp73 suppresses tumor angiogenesis through repression of proangiogenic cytokines and HIF-1α activity
- PMID: 25535357
- PMCID: PMC4291634
- DOI: 10.1073/pnas.1421697112
TAp73 suppresses tumor angiogenesis through repression of proangiogenic cytokines and HIF-1α activity
Abstract
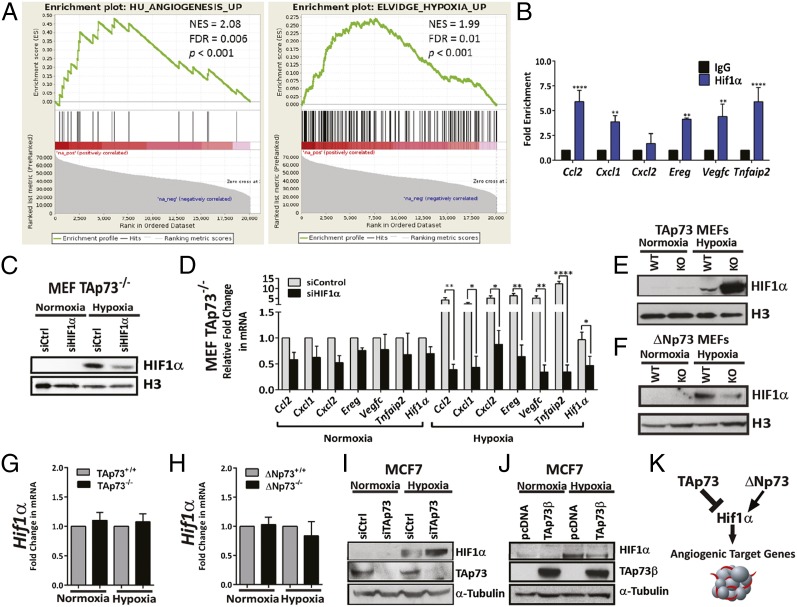
The p53-family member TAp73 is known to function as a tumor suppressor and regulates genomic integrity, cellular proliferation, and apoptosis; however, its role in tumor angiogenesis is poorly understood. Here we demonstrate that TAp73 regulates tumor angiogenesis through repression of proangiogenic and proinflammatory cytokines. Importantly, loss of TAp73 results in highly vascularized tumors, as well as an increase in vessel permeability resulting from disruption of vascular endothelial-cadherin junctions between endothelial cells. In contrast, loss of the oncogenic p73 isoform ΔNp73 leads to reduced blood vessel formation in tumors. Furthermore, we show that up-regulated ΔNp73 levels are associated with increased angiogenesis in human breast cancer and that inhibition of TAp73 results in an accumulation of HIF-1α and up-regulation of HIF-1α target genes. Taken together, our data demonstrate that loss of TAp73 or ΔNp73 up-regulation activates the angiogenic switch that stimulates tumor growth and progression.
Keywords: HIF-1 alpha; angiogenesis; p73; tumor microenvironment; vascular permeability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Grob TJ, et al. Human delta Np73 regulates a dominant negative feedback loop for TAp73 and p53. Cell Death Differ. 2001;8(12):1213–1223. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
